Chapitre 3.4
De la lumière au message nerveux
OBJECTIF
I. Le cristallin est une lentille vivante
• A. Organisation du globe oculaire
TP 1. Rôles des structures internes de l'œil
• B. Le cristallin est formé de cellules transparentes
• C. Le cristallin permet l'accomodation
• D. Anomalies du cristallin et défauts de vision
• L'essentiel
II. La rétine est une surface photosensible
• A. La rétine est formée de trois couches principales de neurones
• B. Les photorécepteurs sont des cônes et des bâtonnets
• C. Les photorécepteurs ne sont pas répartis uniformément
• D. Une chaîne neuronique permet la naissance du message nerveux
• E. Anomalies des pigments rétiniens et vision des couleurs
• L'essentiel
TP 2. Comparaisons moléculaires des opsines
III. Les photorécepteurs sont des produits de l'évolution
• A. Les gènes des opsines : une famille multigénique
• B. L’étude des gènes des opsines permet de placer l’Homme parmi les primates
• L'essentiel
BILAN
I. Le cristallin est une lentille vivante
• A. Organisation du globe oculaire
TP 1. Rôles des structures internes de l'œil
• B. Le cristallin est formé de cellules transparentes
• C. Le cristallin permet l'accomodation
• D. Anomalies du cristallin et défauts de vision
• L'essentiel
II. La rétine est une surface photosensible
• A. La rétine est formée de trois couches principales de neurones
• B. Les photorécepteurs sont des cônes et des bâtonnets
• C. Les photorécepteurs ne sont pas répartis uniformément
• D. Une chaîne neuronique permet la naissance du message nerveux
• E. Anomalies des pigments rétiniens et vision des couleurs
• L'essentiel
TP 2. Comparaisons moléculaires des opsines
III. Les photorécepteurs sont des produits de l'évolution
• A. Les gènes des opsines : une famille multigénique
• B. L’étude des gènes des opsines permet de placer l’Homme parmi les primates
• L'essentiel
BILAN
OBJECTIF
● Nous percevons notre environnement grâce aux organes sensoriels. Parmi eux, l'œil est spécialisé dans la vision. Celui-ci est sensible aux stimulations lumineuses et il élabore un message nerveux conduit jusqu'au cerveau.
● On cherche à préciser comment le stimulus lumineux est perçu ainsi que les implications de ces connaissances en matière de santé.
● Nous percevons notre environnement grâce aux organes sensoriels. Parmi eux, l'œil est spécialisé dans la vision. Celui-ci est sensible aux stimulations lumineuses et il élabore un message nerveux conduit jusqu'au cerveau.
● On cherche à préciser comment le stimulus lumineux est perçu ainsi que les implications de ces connaissances en matière de santé.
1  2
2  3
3  4
4  5
5 
Dissection de l' œil de veau
1. Œil gauche, côté temporal à gauche, côté nasal à droite (la cornée est opaque car l'oeil a été conervé dans du formol) - 2. Hémisphère postérieur - 3. Hémisphère antérieur - 4. Hémisphère antérieur humeur vitrée enlevée - 5. Hémisphère antérieur cristallin enlevé.
Images : www.snv.jussieu.fr
 2
2 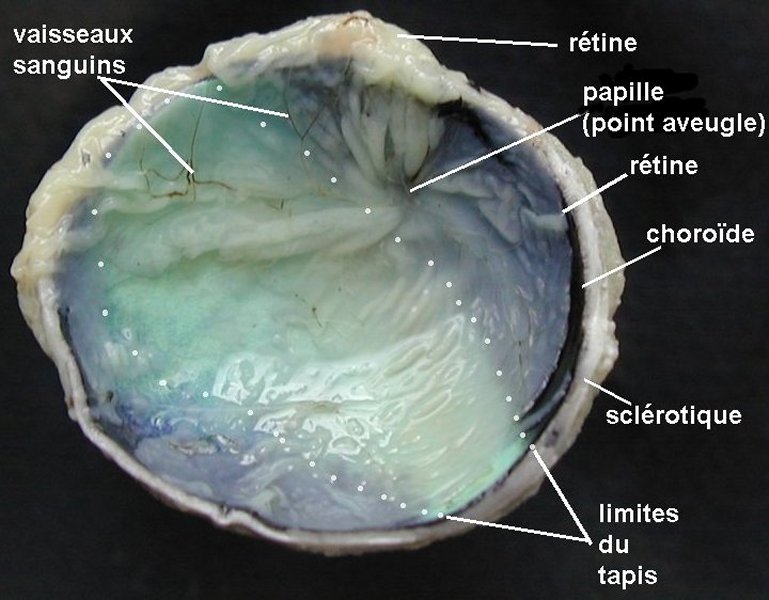 3
3  4
4  5
5 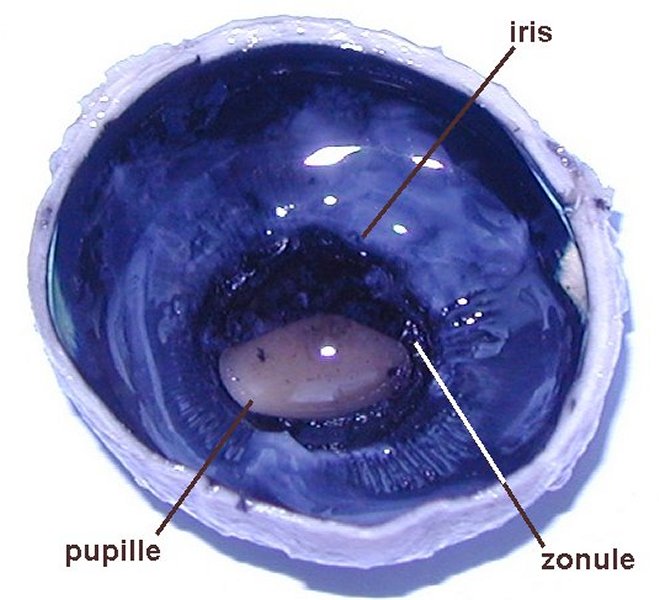
Dissection de l' œil de veau
1. Œil gauche, côté temporal à gauche, côté nasal à droite (la cornée est opaque car l'oeil a été conervé dans du formol) - 2. Hémisphère postérieur - 3. Hémisphère antérieur - 4. Hémisphère antérieur humeur vitrée enlevée - 5. Hémisphère antérieur cristallin enlevé.
Images : www.snv.jussieu.fr
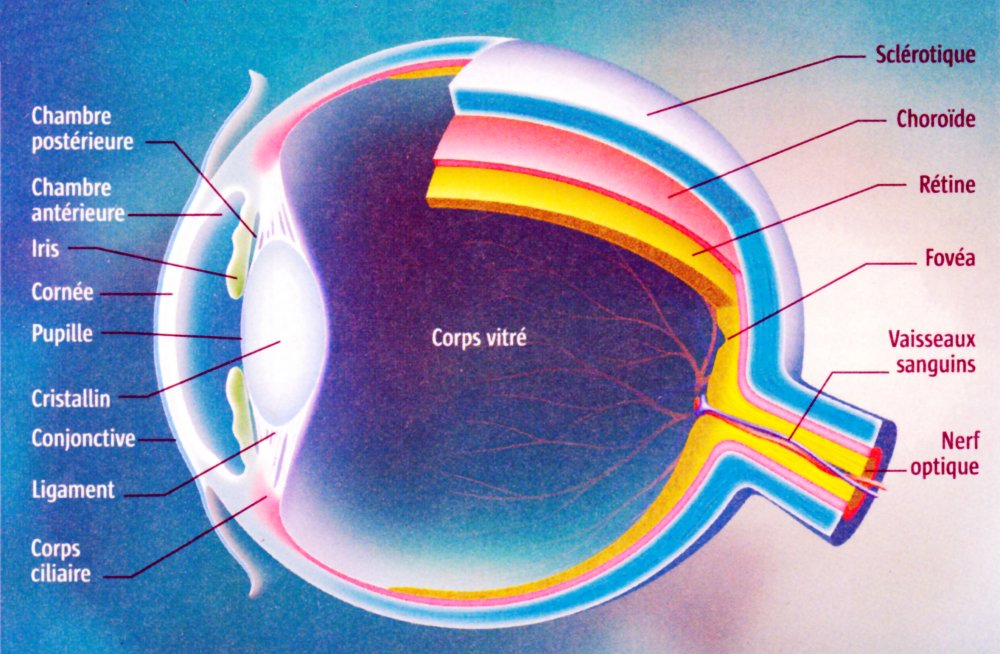
L'œil humain
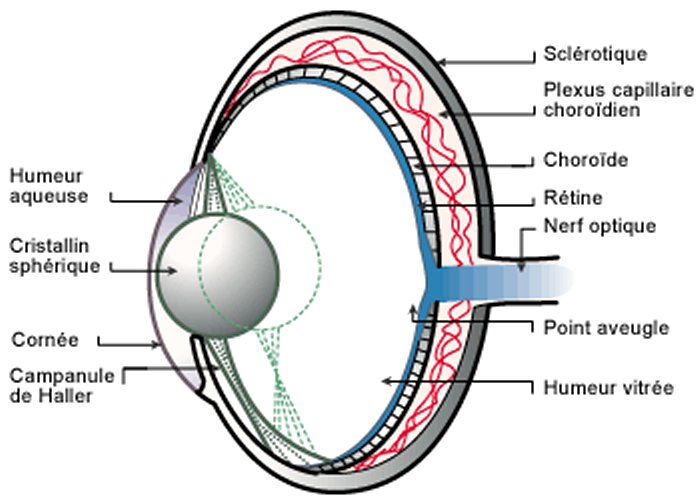
L'œil est un organe quasi sphérique. La conjonctive est une membrane transparente qui produit du mucus et lubrifie la face antérieure de l'œil. La sclérotique forme le globe oculaire, blanche et opaque elle se poursuit vers l'avant par la cornée transparente. Au dessous, la choroïde est une enveloppe vascularisée qui maintient l’œil en chambre noire. Sa partie antérieure forme l'iris pigmenté, circulaire, contractile et percé en son centre d'un orifice de diamètre variable, la pupille. En contrôlant la quantité de lumière qui entre dans l'œil l'iris joue un rôle de diaphragme. L’humeur aqueuse est un liquide transparent constitué à plus de 99% d'eau et qui remplit l'espace situé entre le cristallin et la cornée. L'humeur vitrée est un gel transparent, formé de 95 % d'eau et qui maintient la rétine en place contre la paroi de l’œil. La pression exercée par l'humeur aqueuse et l'humeur vitrée permet le maintien de la forme du globe oculaire. La nature et le rôle du cristallin et de la rétine sont précisés ci-après.
Image : La Recherche n°373 - mars 2004 p. 62 - Voir aussi : www.lyceepmf-tunis.com
L'œil est un organe quasi sphérique. La conjonctive est une membrane transparente qui produit du mucus et lubrifie la face antérieure de l'œil. La sclérotique forme le globe oculaire, blanche et opaque elle se poursuit vers l'avant par la cornée transparente. Au dessous, la choroïde est une enveloppe vascularisée qui maintient l’œil en chambre noire. Sa partie antérieure forme l'iris pigmenté, circulaire, contractile et percé en son centre d'un orifice de diamètre variable, la pupille. En contrôlant la quantité de lumière qui entre dans l'œil l'iris joue un rôle de diaphragme. L’humeur aqueuse est un liquide transparent constitué à plus de 99% d'eau et qui remplit l'espace situé entre le cristallin et la cornée. L'humeur vitrée est un gel transparent, formé de 95 % d'eau et qui maintient la rétine en place contre la paroi de l’œil. La pression exercée par l'humeur aqueuse et l'humeur vitrée permet le maintien de la forme du globe oculaire. La nature et le rôle du cristallin et de la rétine sont précisés ci-après.
Image : La Recherche n°373 - mars 2004 p. 62 - Voir aussi : www.lyceepmf-tunis.com

Le cristallin
Avant d'atteindre la rétine, les rayons lumineux doivent d'abord traverser quatre milieux transparents : la cornée (une dizaine de couches de collagène* contenant très peu de cellules), l'humeur vitrée et l'humeur aqueuse (essentiellement composées d'eau) ainsi que le cristallin formé d'un millier de couches cellulaires empilées (sur 5 mm). Pourtant, ils ne sont ni dispersés, ni réfléchis, ni absorbés. Cela s'explique par l'absence de vascularisation et par un contenu cellulaire transparent et parfaitement homogène.
Avant d'atteindre la rétine, les rayons lumineux doivent d'abord traverser quatre milieux transparents : la cornée (une dizaine de couches de collagène* contenant très peu de cellules), l'humeur vitrée et l'humeur aqueuse (essentiellement composées d'eau) ainsi que le cristallin formé d'un millier de couches cellulaires empilées (sur 5 mm). Pourtant, ils ne sont ni dispersés, ni réfléchis, ni absorbés. Cela s'explique par l'absence de vascularisation et par un contenu cellulaire transparent et parfaitement homogène.
* Le collagène est "une" protéine structurale inextensible, particulièrement abondante dans l'organisme et présente dans la matrice intercellulaire.
Voir : fr.wikipedia.org
Voir : fr.wikipedia.org
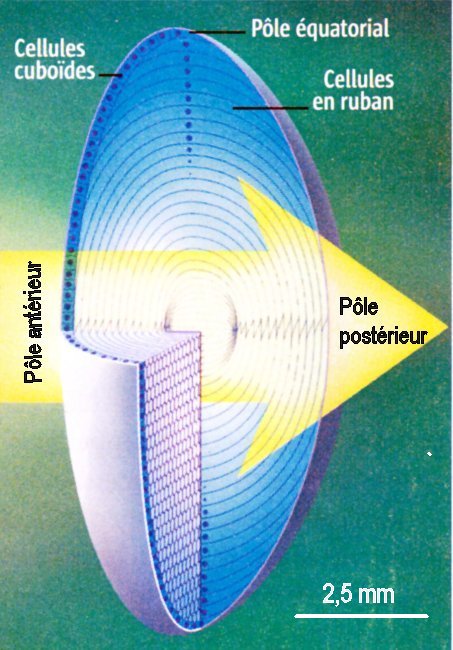
1 2
2 
1. Organisation générale du cristallin 2. Cellules du cristallin (MO x 100)
Le cristallin est enveloppé par une membrane acellulaire (2a). Au dessous, et sur la face antérieure, se trouve une monocouche de cellules cuboïdes (2b) qui se divisent chez le jeune individu et repoussent vers l'intérieur les cellules plus anciennes (migration). Durant cette migration à partir du pôle équatorial, les cellules s'allongent jusqu'à atteindre les pôles antérieur et postérieur du cristallin. Ces cellules en ruban (ou en lames) perdent ensuite leur noyau et leurs organites de sorte que, dans la partie centrale du cristallin, leur cytoplame est constitué à plus de 90% de protéines appelées cristallines qui forment un gel optiquement très homogène et totalement transparent. Les cellules du cristallin ne sont jamais remplacées, mais de nouvelles cellules se forment et s'ajoutent en périphérie (les divisions, qui débutent chez l'embryon, cessent vers l'âge de 20 ans).
Images (modifiées) : 1 La Recherche n°373 - mars 2004 p. 62 - 2 www.larousse.fr
 2
2 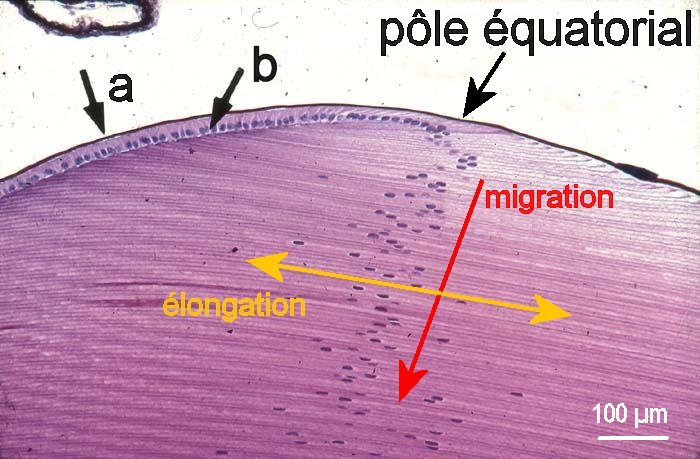
1. Organisation générale du cristallin 2. Cellules du cristallin (MO x 100)
Le cristallin est enveloppé par une membrane acellulaire (2a). Au dessous, et sur la face antérieure, se trouve une monocouche de cellules cuboïdes (2b) qui se divisent chez le jeune individu et repoussent vers l'intérieur les cellules plus anciennes (migration). Durant cette migration à partir du pôle équatorial, les cellules s'allongent jusqu'à atteindre les pôles antérieur et postérieur du cristallin. Ces cellules en ruban (ou en lames) perdent ensuite leur noyau et leurs organites de sorte que, dans la partie centrale du cristallin, leur cytoplame est constitué à plus de 90% de protéines appelées cristallines qui forment un gel optiquement très homogène et totalement transparent. Les cellules du cristallin ne sont jamais remplacées, mais de nouvelles cellules se forment et s'ajoutent en périphérie (les divisions, qui débutent chez l'embryon, cessent vers l'âge de 20 ans).
Images (modifiées) : 1 La Recherche n°373 - mars 2004 p. 62 - 2 www.larousse.fr

Cellules en ruban du cristallin (MEB). Dépourvues d'espace intercellulaire (l'écart entre deux cellules est inférieur à la longueur d'onde de la lumière visible), elles forment des lames à section hexagonale d'1 cm de long pour 10 µm de large et 4,5 µm d'épaisseur. La lumière arrive perpendiculairement à leur surface, ce qui évite sa dispersion.
Image (modifiée) : www.larousse.fr
Image (modifiée) : www.larousse.fr
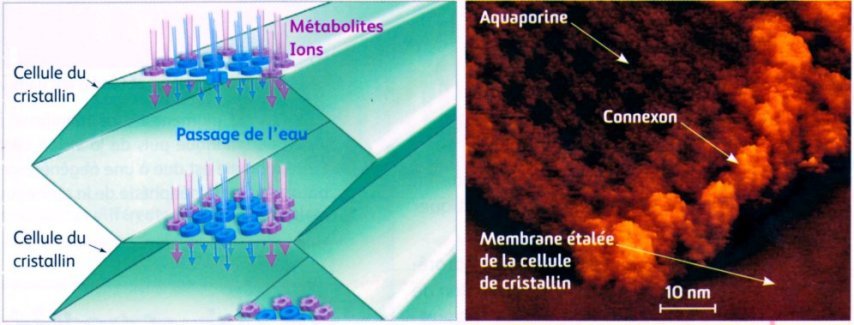
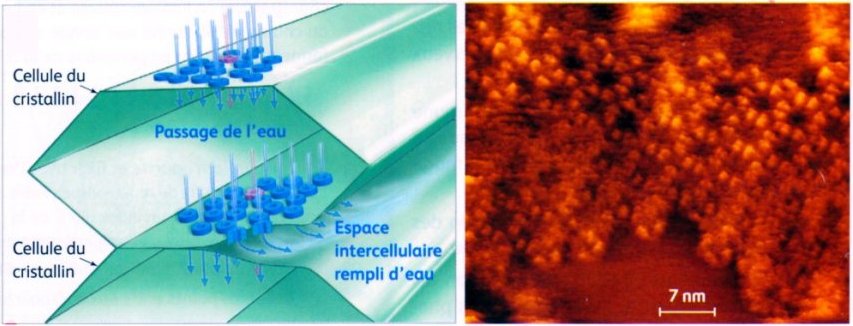
Membrane plamique des cellules du cristallin (AFM) (la microscopie à force atomique (AFM) est une technique qui permet d’obtenir une image de la surface d’un échantillon avec une précision d’un milliardième de mètre). Privées de vascularisation et d'organites, les cellules du cristallin n'en ont pas moins une longévité exceptionnelle, elles peuvent rester fonctionnelles une centaine d'année sans jamais être supprimées ni remplacées. Pour assurer leurs échanges métaboliques (eau, ions, glucose, déchets) avec les humeurs vitrée et aqueuse elles dépendent entièrement de deux types de canaux qui assurent aussi l'adhésion cellulaire : les aquaporines qui permettent le passage d'eau et les connexons qui permettent le passage des métabolites et des ions.
Image (modifiée) : SVT 1S, Nathan 2011 p. 349 - Source : www2.cnrs.fr
Image (modifiée) : SVT 1S, Nathan 2011 p. 349 - Source : www2.cnrs.fr
Rappels de physique
Dans un milieu transparent et homogène la lumière se propage en ligne droite. Cependant, quand un rayon lumineux passe d'un milieu transparent dans un autre, il change de vitesse et subit une déviation, c'est la réfraction. L'angle de réfraction dépend de l'angle d'incidence et de l'indice de réfraction des milieux traversés.
Dans un milieu transparent et homogène la lumière se propage en ligne droite. Cependant, quand un rayon lumineux passe d'un milieu transparent dans un autre, il change de vitesse et subit une déviation, c'est la réfraction. L'angle de réfraction dépend de l'angle d'incidence et de l'indice de réfraction des milieux traversés.

Le cristallin a les propriétés d'une lentille convergente
Le cristallin a la forme d'une lentille biconvexe. Ses cellules contiennent principalement de la cristalline qui a un indice de réfraction quasi identique à celui de la membrane cellulaire mais supérieur à celui de l'eau de l'humeur aqueuse et de l'humeur vitrée qui l'entourent. Les rayons lumineux qui traversent le cristallin subissent donc une double réfraction qui entraîne leur convergence vers l'axe optique de l'œil (en un point F').
Le cristallin a la forme d'une lentille biconvexe. Ses cellules contiennent principalement de la cristalline qui a un indice de réfraction quasi identique à celui de la membrane cellulaire mais supérieur à celui de l'eau de l'humeur aqueuse et de l'humeur vitrée qui l'entourent. Les rayons lumineux qui traversent le cristallin subissent donc une double réfraction qui entraîne leur convergence vers l'axe optique de l'œil (en un point F').
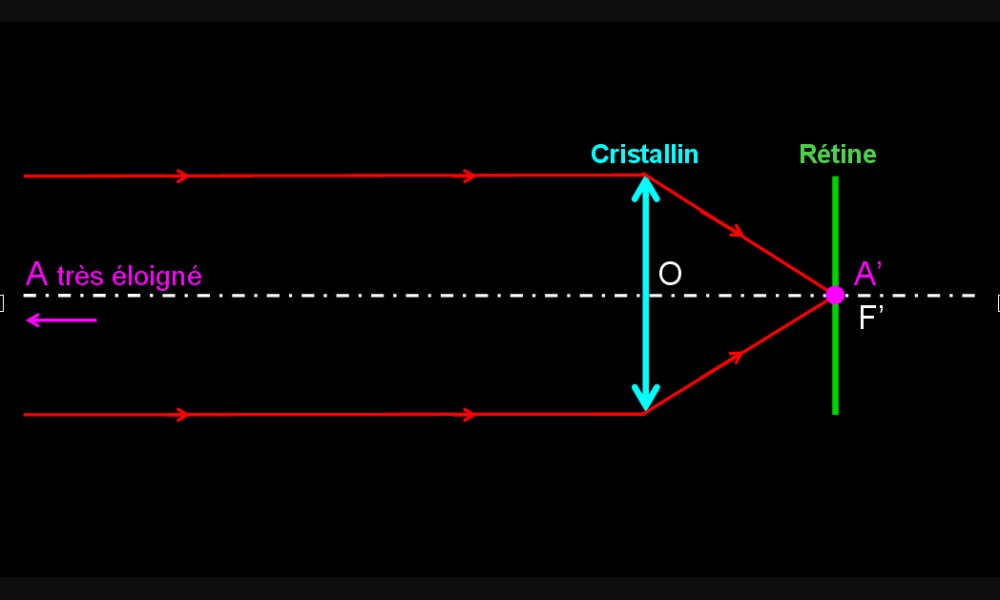
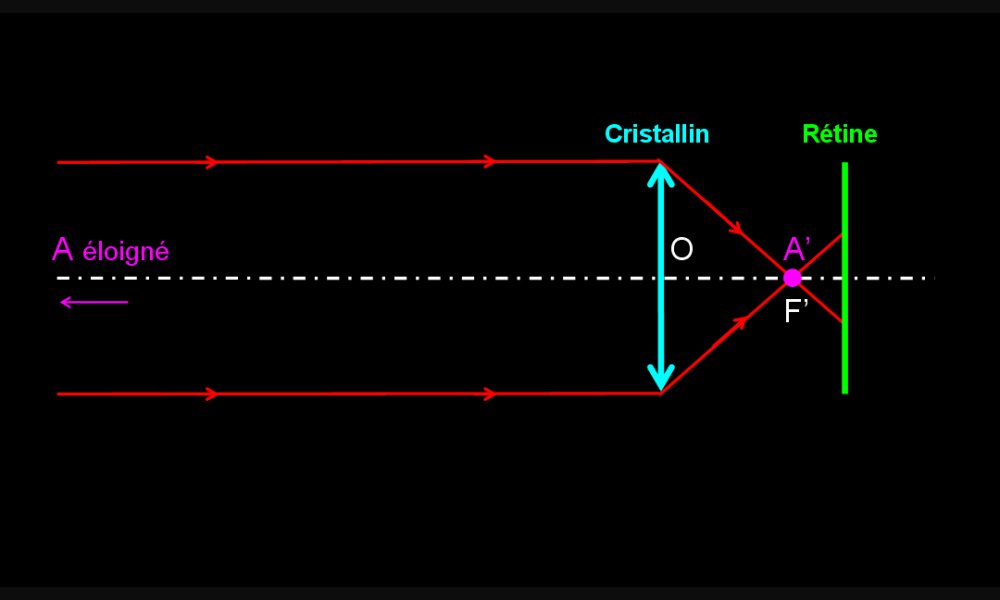
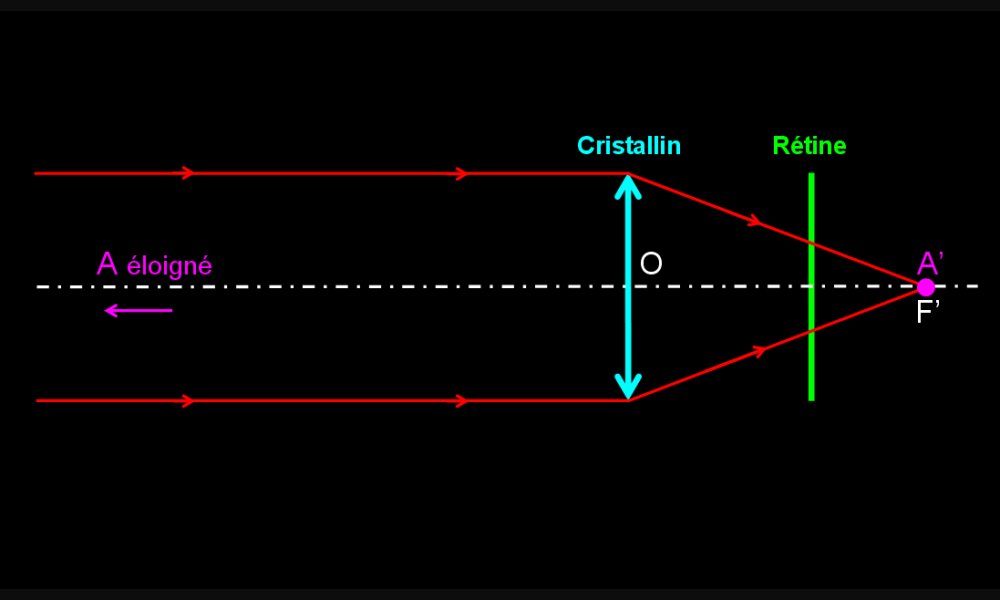
Formation de l’image d’un objet lointain par un œil emmétrope
Les rayons lumineux qui nous parviennent d'un objet lointain (à l'infini) sont parallèles. L'objet (A) est vu nettement car son image (A') se forme au foyer image (F') qui est situé sur la rétine. Un rayon passant par le centre optique (O) n'est pas dévié.
Le foyer image (F') d'un lentille est le point de l'axe optique vers lequel les rayons incidents parallèles à l'axe optique, sont déviés.
La distance focale d'une lentille est la distance séparant la lentille du foyer image.
Les rayons lumineux qui nous parviennent d'un objet lointain (à l'infini) sont parallèles. L'objet (A) est vu nettement car son image (A') se forme au foyer image (F') qui est situé sur la rétine. Un rayon passant par le centre optique (O) n'est pas dévié.
Le foyer image (F') d'un lentille est le point de l'axe optique vers lequel les rayons incidents parallèles à l'axe optique, sont déviés.
La distance focale d'une lentille est la distance séparant la lentille du foyer image.
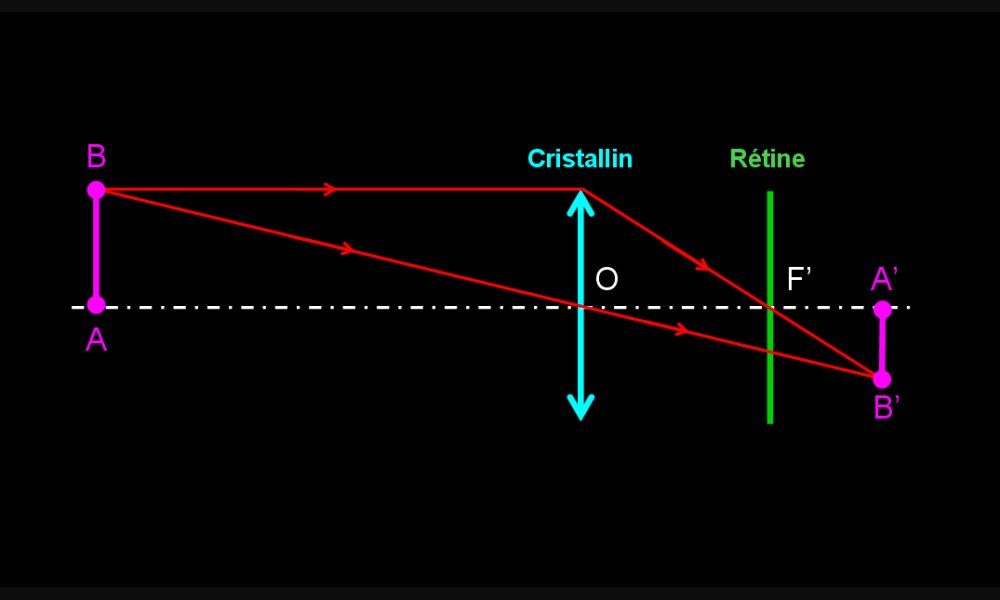
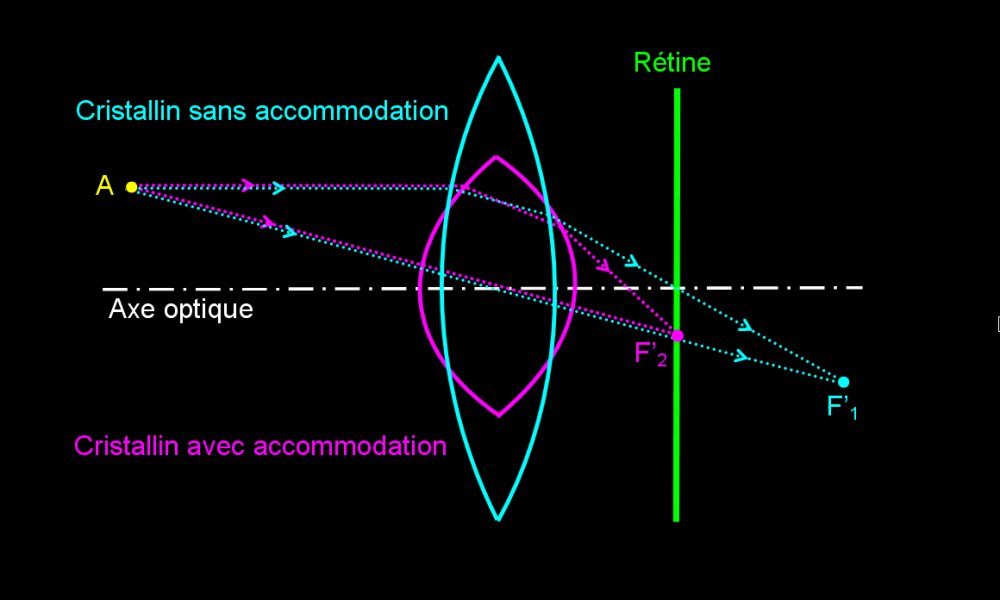
Formation de l’image d’un objet proche par un œil emmétrope sans accommodation
Les rayons lumineux qui nous parviennent d'un objet proche (AB) sont obliques. L'image (A' B') a tendance à se former en arrière de la rétine. Elle serait donc floue sans modification des propriétés de l'œil.
Les rayons lumineux qui nous parviennent d'un objet proche (AB) sont obliques. L'image (A' B') a tendance à se former en arrière de la rétine. Elle serait donc floue sans modification des propriétés de l'œil.

Formation de l’image d’un objet proche par un œil emmétrope avec accommodation
Le diamètre du globe oculaire étant constant, chez vertébrés terrestres c'est la convergence du cristallin (= vergence) qui augmente. Cela a pour effet de raccourcir la distance focale et de permettre à l'image (A' B') de se former sur la rétine. C'est l'accomodation.
L'œil a une distance focale variable qui permet de former sur la rétine l'image d'objets situés à des distances variables.
Le diamètre du globe oculaire étant constant, chez vertébrés terrestres c'est la convergence du cristallin (= vergence) qui augmente. Cela a pour effet de raccourcir la distance focale et de permettre à l'image (A' B') de se former sur la rétine. C'est l'accomodation.
L'œil a une distance focale variable qui permet de former sur la rétine l'image d'objets situés à des distances variables.
Animations numériques
Lentilles minces (lien externe) : la convergence de la lumière et Types de lentilles (lien externe) : lentille biconvexe
Source : ressources.univ-lemans.fr
Lentilles minces (lien externe) : la convergence de la lumière et Types de lentilles (lien externe) : lentille biconvexe
Source : ressources.univ-lemans.fr

Une image inversée
L'image obtenue sur la rétine est renversée de haut en bas et inversée de gauche à droite. C'est le cerveau qui interprète les images reçues par la rétine et qui nous rend un monde extérieur "à l'endroit".
Image (modifiée) : perso.id-net.fr
L'image obtenue sur la rétine est renversée de haut en bas et inversée de gauche à droite. C'est le cerveau qui interprète les images reçues par la rétine et qui nous rend un monde extérieur "à l'endroit".
Image (modifiée) : perso.id-net.fr
![]()

Insertion des muscles ciliaires sur le cristallin
Les muscles ciliaires (situés dans corps ciliaire qui est la partie antérieure différenciée de la chroroïde) forment un anneau relié au cristallin par les ligaments suspenseurs (= zonule).
Image : SVT 1eS, Belin 2011, p. 293
{kind=link}
Les muscles ciliaires (situés dans corps ciliaire qui est la partie antérieure différenciée de la chroroïde) forment un anneau relié au cristallin par les ligaments suspenseurs (= zonule).
Image : SVT 1eS, Belin 2011, p. 293
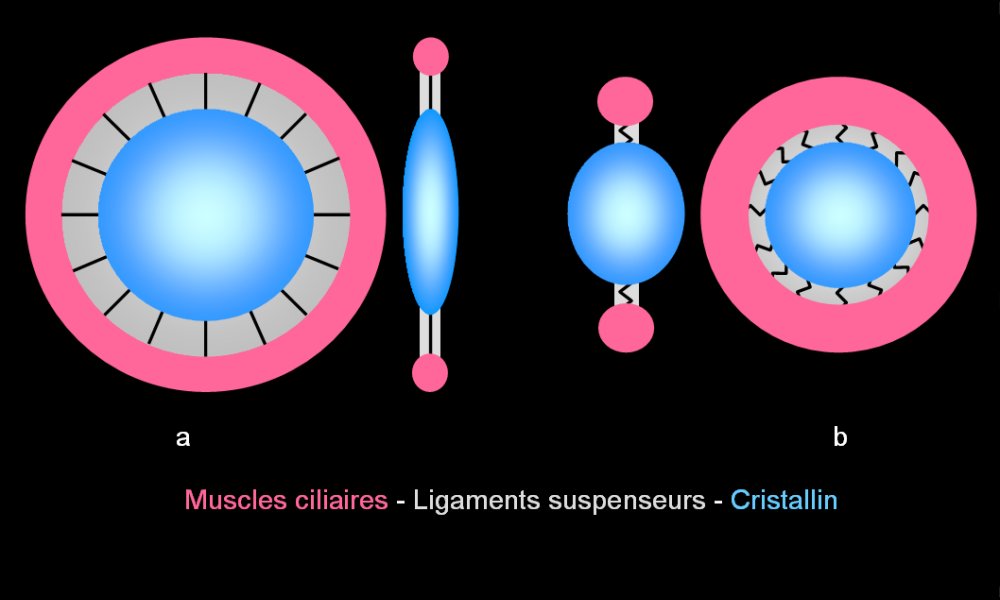
Modification du diamètre du cristallin
a. Quand les muscles ciliaires sont relâchés, leur diamètre est maximum. Les ligaments suspenseurs sont tendus et exercent des tractions radiales sur le cristallin qui a alors un diamètre maximum et prend une forme applatie.
b. Quand les muscles ciliaires sont contractés, leur diamètre est réduit. Les ligaments suspenseurs sont détendus. Le cristallin diminue de diamètre et prend spontanément une forme bombée grâce à son élasticité.
a. Quand les muscles ciliaires sont relâchés, leur diamètre est maximum. Les ligaments suspenseurs sont tendus et exercent des tractions radiales sur le cristallin qui a alors un diamètre maximum et prend une forme applatie.
b. Quand les muscles ciliaires sont contractés, leur diamètre est réduit. Les ligaments suspenseurs sont détendus. Le cristallin diminue de diamètre et prend spontanément une forme bombée grâce à son élasticité.
Modification de l'épaisseur du cristallin
La distance focale de l'œil est maximale (17 mm pour l'œil humain) quand les muscles ciliaires sont au repos. Dans ces conditions l'œil n'accommode pas et on peut voir des objets situés à une distance maximale appelée punctum remotum, qui est infinie pour un œil normal (œil emmetrope).
La distance focale de l'œil est minimale quand les muscles ciliaires sont contractés, l'œil accommode. On peut alors voir nettement des objets situés à une distance minimale appelée punctum proximum, qui est d'environ 25 cm pour un œil émmétrope moyen.
Remarque. On parle de "distance focale de l'œil" et non de celle du cristallin car l'œil est un système optique formé de plusieurs milieux convergents (cornée, humeur aqueuse, cristallin, humeur vitrée). En revanche l'accomodation dépend uniquement du cristallin.
La distance focale de l'œil est maximale (17 mm pour l'œil humain) quand les muscles ciliaires sont au repos. Dans ces conditions l'œil n'accommode pas et on peut voir des objets situés à une distance maximale appelée punctum remotum, qui est infinie pour un œil normal (œil emmetrope).
La distance focale de l'œil est minimale quand les muscles ciliaires sont contractés, l'œil accommode. On peut alors voir nettement des objets situés à une distance minimale appelée punctum proximum, qui est d'environ 25 cm pour un œil émmétrope moyen.
Remarque. On parle de "distance focale de l'œil" et non de celle du cristallin car l'œil est un système optique formé de plusieurs milieux convergents (cornée, humeur aqueuse, cristallin, humeur vitrée). En revanche l'accomodation dépend uniquement du cristallin.

Le punctum proximum est faible chez les enfants et s'accroît régulièrement avec l'âge.
L'accomodation chez les poissons
Chez les poissons le cristallin ne change pas de forme et reste constamment sphérique. L'accomodauion se fait par déplacement du cristallin et non pas par déformation.
Image www.jeulin.fr
Chez les poissons le cristallin ne change pas de forme et reste constamment sphérique. L'accomodauion se fait par déplacement du cristallin et non pas par déformation.
Image www.jeulin.fr
La myopie
Si le cristallin est trop convergent (distance focale au repos trop courte), l'image d'un objet à l'infini se forme avant la rétine. La vision de loin est perturbée.
Remarque. L'œil myope peut être aussi trop profond avec un cristallin normal. Cela donne le même résultat.
Si le cristallin est trop convergent (distance focale au repos trop courte), l'image d'un objet à l'infini se forme avant la rétine. La vision de loin est perturbée.
Remarque. L'œil myope peut être aussi trop profond avec un cristallin normal. Cela donne le même résultat.
L'hypermétropie
Si le cristallin n'est pas assez convergent (distance focale au repos trop grande), l'image d'un objet à l'infini se forme en arrière de la rétine. L'œil accommmode alors en vision de loin. L'accommodation ayant des limites, la vision de près peut être perturbée.
Remarque. L'œil hypermétrope peut être aussi trop peu profond avec un cristallin normal. Cela donne le même résultat.
Si le cristallin n'est pas assez convergent (distance focale au repos trop grande), l'image d'un objet à l'infini se forme en arrière de la rétine. L'œil accommmode alors en vision de loin. L'accommodation ayant des limites, la vision de près peut être perturbée.
Remarque. L'œil hypermétrope peut être aussi trop peu profond avec un cristallin normal. Cela donne le même résultat.
La presbytie
À partir d'environ 45 ans le cristallin perd peu à peu son pouvoir d'accommodation. Le punctum proximum se rapproche du punctum remotum. Le sujet doit progressivement éloigner les textes qu'il lit mais sa vision de loin n'est pas affectée.
Remarque 1. Un myope n'aura pas besoin, pendant un certain temps, de corriger sa vision de près qui était meilleure que la moyenne (par contre sa vision de loin ne s'améliorera pas).
Remarque 2. Dans certains cas, la presbytie peut révéler une hypermétropie passée inaperçu. Une hypermétropie légère pouvant en effet être compensée par un effort d'accommodation. Avec l'âge, la capacité d'accommodation diminuant, l'hypermétropie se révèle. La presbytie est alors précoce.
À partir d'environ 45 ans le cristallin perd peu à peu son pouvoir d'accommodation. Le punctum proximum se rapproche du punctum remotum. Le sujet doit progressivement éloigner les textes qu'il lit mais sa vision de loin n'est pas affectée.
Remarque 1. Un myope n'aura pas besoin, pendant un certain temps, de corriger sa vision de près qui était meilleure que la moyenne (par contre sa vision de loin ne s'améliorera pas).
Remarque 2. Dans certains cas, la presbytie peut révéler une hypermétropie passée inaperçu. Une hypermétropie légère pouvant en effet être compensée par un effort d'accommodation. Avec l'âge, la capacité d'accommodation diminuant, l'hypermétropie se révèle. La presbytie est alors précoce.
1
 2
2 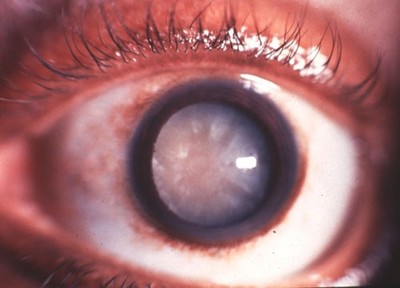 4
4 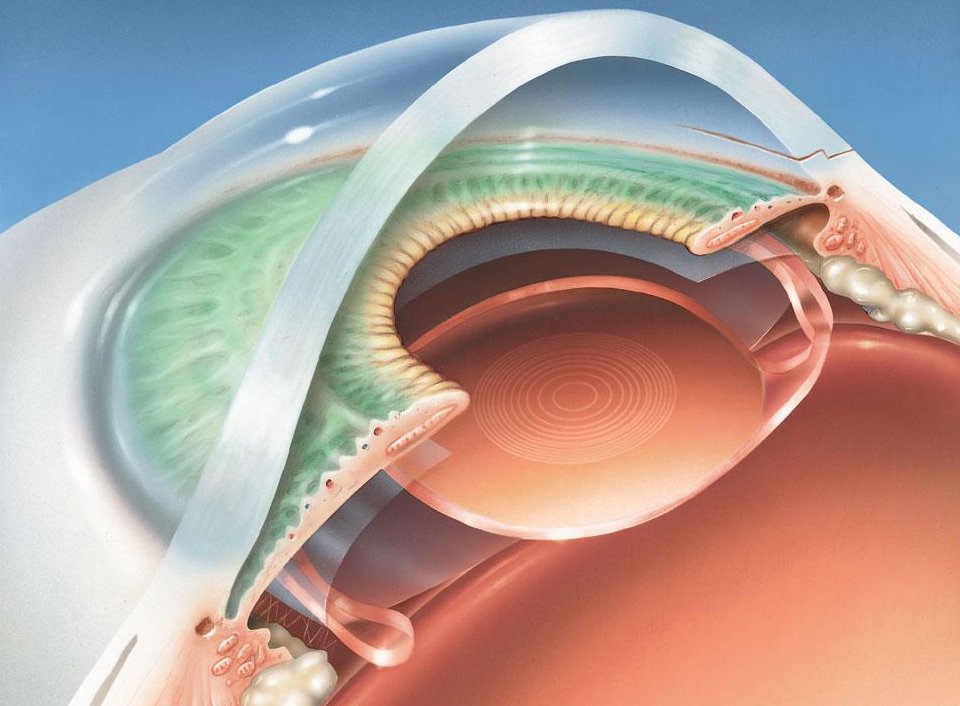
1. Membrane plamique des cellules du cristallin d'un patient atteint de cataracte (micoscopie AFM) - 2 Mesure des cristallines intactes dans des cristallins d'âge différent - 3. Œil d'une personne atteinte de cataracte (forme très avancée) - 4 Cristallin artificiel
Les cellules en ruban ne possèdent plus ni l'information génétique ni les organites permettant de renouveler les cristallines (voir I.B). Au fil des années l'accumulation de micro-lésions ainsi que les dommages causés par les rayons ultraviolets ou par des taux de sucre importants (diabète) sont irréversibles. Les cristallines finissent par précipiter. La perte de connexons (1) entraîne un manque d’adhérence, l’accumulation de déchets dans les cellules et des défauts de transport de l’eau qui envahit les espaces intercellulaires. Les cellules se détachent les unes des autres, la lumière ne traverse plus librement le cristallin (2). Cela entraîne la cataracte (3) qui atteint plus d'une personne sur cinq à partir de 65 ans, plus d'une sur trois à partir de 75 ans et près de deux sur trois après 85 ans. Elle se caractérise par une baisse de la vue, une impression de brouillard, un éblouissement à la lumière vive.
La cataracte est la cause de près de 40 % des 37 millions d'aveugles dans le monde, c'est la première cause de cécité dans le tiers-monde. Actuellement, le seul traitement efficace de la cataracte est la chirurgie. L'intervention consiste à enlever le cristallin opaque et à le remplacer par un cristallin artificiel (4).
Images : 1 et 2 SVT 1S, Nathan 2011 p. 349 - 3 histoblog.viabloga.com - 4 www.centrevisionbretagne.com
Les cellules en ruban ne possèdent plus ni l'information génétique ni les organites permettant de renouveler les cristallines (voir I.B). Au fil des années l'accumulation de micro-lésions ainsi que les dommages causés par les rayons ultraviolets ou par des taux de sucre importants (diabète) sont irréversibles. Les cristallines finissent par précipiter. La perte de connexons (1) entraîne un manque d’adhérence, l’accumulation de déchets dans les cellules et des défauts de transport de l’eau qui envahit les espaces intercellulaires. Les cellules se détachent les unes des autres, la lumière ne traverse plus librement le cristallin (2). Cela entraîne la cataracte (3) qui atteint plus d'une personne sur cinq à partir de 65 ans, plus d'une sur trois à partir de 75 ans et près de deux sur trois après 85 ans. Elle se caractérise par une baisse de la vue, une impression de brouillard, un éblouissement à la lumière vive.
Images : 1 et 2 SVT 1S, Nathan 2011 p. 349 - 3 histoblog.viabloga.com - 4 www.centrevisionbretagne.com

Claude Monet et la cataracte
Claude Monet (1840-1926) est un peinte impresionniste (lien externe). En 1882 il s'installe à Giverny où il crée, en 1890, un jardin d'eau avec le bassin aux nymphéas qu'il peint plusieurs fois à partir de 1897. À gauche, Le bassin aux Nymphéas, peint en 1899 avant la maladie, à droite, Le pont japonais, peint en 1923 quand les deux yeux du l'artiste sont atteints par la cataracte. Les teintes s'accentuent dans les rouges et les jaunes, les bleus ont tendance à disparaître. Les détails s'estompent également.
Images : www.intermonet.com
Claude Monet (1840-1926) est un peinte impresionniste (lien externe). En 1882 il s'installe à Giverny où il crée, en 1890, un jardin d'eau avec le bassin aux nymphéas qu'il peint plusieurs fois à partir de 1897. À gauche, Le bassin aux Nymphéas, peint en 1899 avant la maladie, à droite, Le pont japonais, peint en 1923 quand les deux yeux du l'artiste sont atteints par la cataracte. Les teintes s'accentuent dans les rouges et les jaunes, les bleus ont tendance à disparaître. Les détails s'estompent également.
Images : www.intermonet.com
● Dans l'œil, les rayons lumineux traversent successivement quatre systèmes dépourvus de vascularisation et transparents : la cornée, l'humeur aqueuse, le cristallin et l'humeur vitrée. Ils ne sont ni dispersés, ni réfléchis, ni absorbés.
● Le cristallin est souple et biconvexe. Il est formé de cellules vivantes, sans espace intercellulaire, allongées, aplaties, qui ont perdu leur noyau et leurs organites : les cellules en ruban dont le cytoplasme est essentiellement constitué de protéines (cristallines) qui forment un gel optiquement homogène et transparent. Ces particularités confèrent au cristallin les propriétés d'une lentille convergente à vergence variable grâce à l'action des muscles ciliaires qui modifient sa courbure et permettent l'accommodation (passage de la vision de près à la vision de loin et inversement).
● Le maintien de la transparence des cellules du cristallin nécessite le renouvellement permanent de leur contenu. Des canaux intercellulaires permettent les échanges métaboliques (eau, ions, glucose, déchets) avec les humeurs vitrée et aqueuse ainsi que l'adhésion cellulaire.
● Si le cristallin est trop convergent l'image d'un objet à l'infini se forme en avant de la rétine. La vision de loin est perturbée, c'est la myopie. Si le cristallin n'est pas assez convergent l'image d'un objet à l'infini se forme en arrière de la rétine. La vision de près est perturbée quand l'accommodation ne compense plus l'hypermétropie.
● Les cellules du cristallin ont une longévité exceptionnelle (une centaine d'années sans être remplacées). Avec l'âge leurs propriétés s'altèrent :
- la souplesse du cristallin diminue, ce qui entraîne une difficulté d'accommodation et donc une altération de la vision de près, c'est la presbytie ;
- la transparence du cristallin diminue, car le renouvellement du contenu cellulaire est perturbé, ce qui entraîne une altération de la perception de la luminosité et des couleurs, c'est la cataracte.
● Le cristallin est souple et biconvexe. Il est formé de cellules vivantes, sans espace intercellulaire, allongées, aplaties, qui ont perdu leur noyau et leurs organites : les cellules en ruban dont le cytoplasme est essentiellement constitué de protéines (cristallines) qui forment un gel optiquement homogène et transparent. Ces particularités confèrent au cristallin les propriétés d'une lentille convergente à vergence variable grâce à l'action des muscles ciliaires qui modifient sa courbure et permettent l'accommodation (passage de la vision de près à la vision de loin et inversement).
● Le maintien de la transparence des cellules du cristallin nécessite le renouvellement permanent de leur contenu. Des canaux intercellulaires permettent les échanges métaboliques (eau, ions, glucose, déchets) avec les humeurs vitrée et aqueuse ainsi que l'adhésion cellulaire.
● Si le cristallin est trop convergent l'image d'un objet à l'infini se forme en avant de la rétine. La vision de loin est perturbée, c'est la myopie. Si le cristallin n'est pas assez convergent l'image d'un objet à l'infini se forme en arrière de la rétine. La vision de près est perturbée quand l'accommodation ne compense plus l'hypermétropie.
● Les cellules du cristallin ont une longévité exceptionnelle (une centaine d'années sans être remplacées). Avec l'âge leurs propriétés s'altèrent :
- la souplesse du cristallin diminue, ce qui entraîne une difficulté d'accommodation et donc une altération de la vision de près, c'est la presbytie ;
- la transparence du cristallin diminue, car le renouvellement du contenu cellulaire est perturbé, ce qui entraîne une altération de la perception de la luminosité et des couleurs, c'est la cataracte.
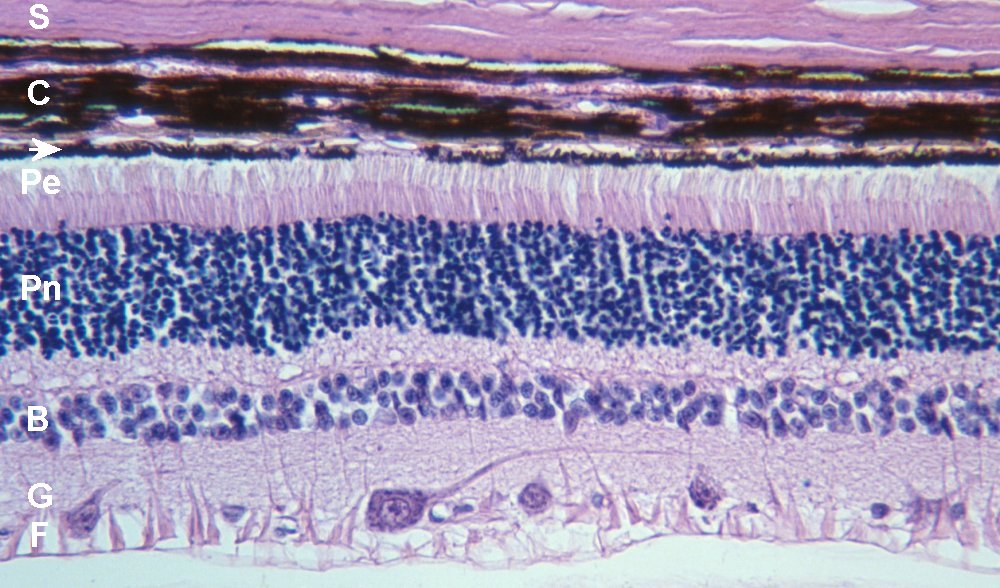
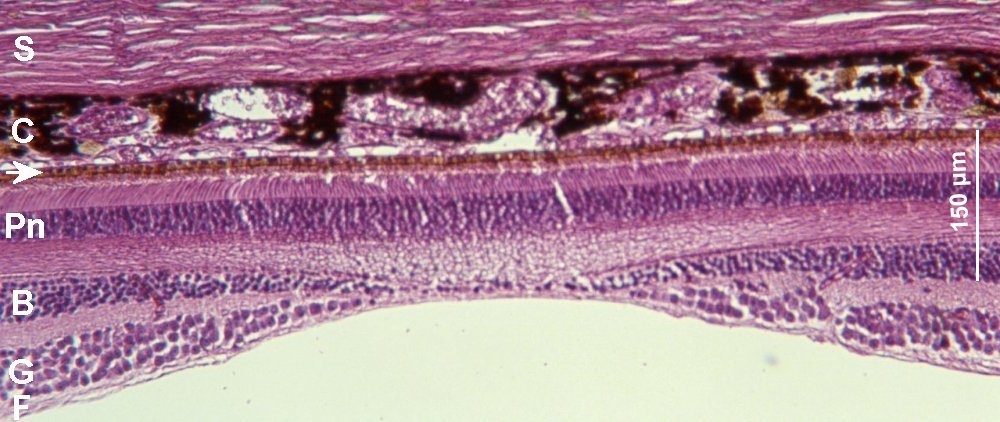
Coupe de rétine (MP x 240) (chien)
S sclérotique ; C choroïde ; T tapetum (pas chez l'Homme) ; flèche épithélium (pigmenté chez l'Homme) ; Pe phorécepteurs (segments externes) ; Pn noyaux des photorécepteurs ; B noyaux des neurones bipolaires (auxquels sont associés les noyaux des cellules horizontales, côté interne, et amacrines, côté externe, qui apparaissent plus gros) ; G cellules ganglionnaires ; F fibres nerveuses.
Les vertébrés ont une rétine inversée. Les rayons lumineux qui traversent l'œil doivent donc traverser la couche de fibres nerveuses et plusieurs couches cellulaires avant d'atteindre les photorécepteurs (l'épaisseur de la rétine est de 400 µm). Cela est possible car la rétine est transparente.
Image : www.vetmed.ucdavis.edu
S sclérotique ; C choroïde ; T tapetum (pas chez l'Homme) ; flèche épithélium (pigmenté chez l'Homme) ; Pe phorécepteurs (segments externes) ; Pn noyaux des photorécepteurs ; B noyaux des neurones bipolaires (auxquels sont associés les noyaux des cellules horizontales, côté interne, et amacrines, côté externe, qui apparaissent plus gros) ; G cellules ganglionnaires ; F fibres nerveuses.
Les vertébrés ont une rétine inversée. Les rayons lumineux qui traversent l'œil doivent donc traverser la couche de fibres nerveuses et plusieurs couches cellulaires avant d'atteindre les photorécepteurs (l'épaisseur de la rétine est de 400 µm). Cela est possible car la rétine est transparente.
Image : www.vetmed.ucdavis.edu
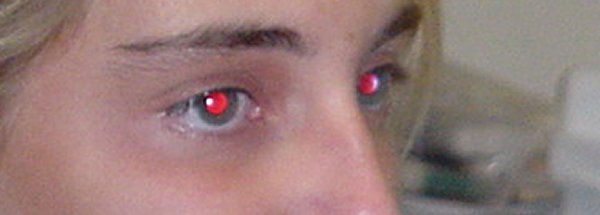
Effet yeux rouges. En milieu peu éclairé la pupille est dilatée. Une photographie prise de face et au flash, permet alors de voir le fond de l'œil. La rétine étant transparente on observe la choroïde qui est très vascularisée.
Image (modifiée) : ecfa.unige.ch
Image (modifiée) : ecfa.unige.ch

Effet yeux verts. Le même effet s'observe aussi chez les autres mammifères mais la couleur n'est pas nécessairement le rouge. La rétine et la choroïde ont pourtant les mêmes propriérés que chez l'Homme mais une couche réfléchissante, le tapis (tapetum) se situe entre la rétine et la choroïde. Les photons qui ne sont pas captés lors du premier passage sont alors réfléchis et peuvent l'être au retour. Ceci permet une meilleure sensibilité dans des conditions de lumière basse. C'est pour cela que les yeux des chats brillent la nuit quand ils sont éclairés. Leur tapetum apparaît vert, mais chez d'autres espèces il est de couleur bleue, jaune ou rose.
Image : fr.wikipedia.org
Image : fr.wikipedia.org
1 2
2 
1. Cônes et bâtonnets (MEB x 6000) et 2. Interprétation
Chaque bâtonnet et chaque cône est formé d'un segment interne, qui contient les organites habituels d'une cellule, et d'un segment externe qui est constitué d'un empilement de membranes formant des disques, dans lesquels est incorporé un pigment rétinien.
Images : 1 www.cnrs.fr - 2 lecerveau.mcgill.ca
 2
2 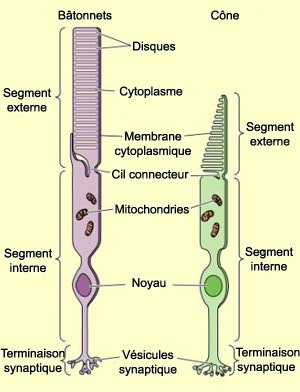
1. Cônes et bâtonnets (MEB x 6000) et 2. Interprétation
Chaque bâtonnet et chaque cône est formé d'un segment interne, qui contient les organites habituels d'une cellule, et d'un segment externe qui est constitué d'un empilement de membranes formant des disques, dans lesquels est incorporé un pigment rétinien.
Images : 1 www.cnrs.fr - 2 lecerveau.mcgill.ca

Représentation moléculaire d'un pigment rétinien
Un pigment rétinien est formé de l'association de deux molécules : l'opsine (protéine transmembranaire) et le rétinal (non protéique et synthétisé à partir de la vitamine A). Quand le rétinal absorbe la lumière sa conformation (sa forme) change ce qui entraîne un changement de celle de l'opsine. Il en résulte une cascade de réactions chimiques conduisant à la transformation du stimulus lumineux en message nerveux conduit par le nerf optique (c'est la phototransduction). Cette situation (état activé) est instable le rétinal et donc l'opsine reprennent spontanément leur conformation d'origine (état passif).
Image : www.psi.ch (modifiée)
Un pigment rétinien est formé de l'association de deux molécules : l'opsine (protéine transmembranaire) et le rétinal (non protéique et synthétisé à partir de la vitamine A). Quand le rétinal absorbe la lumière sa conformation (sa forme) change ce qui entraîne un changement de celle de l'opsine. Il en résulte une cascade de réactions chimiques conduisant à la transformation du stimulus lumineux en message nerveux conduit par le nerf optique (c'est la phototransduction). Cette situation (état activé) est instable le rétinal et donc l'opsine reprennent spontanément leur conformation d'origine (état passif).
Image : www.psi.ch (modifiée)
L'opsine est formée de sept segments transmembranaires (la bande grise représente la membrane cellulaire) entourant le rétinal qui a une forme coudée à l' état passif. Sous l'effet de la lumière le rétinal adopte une conformation rectiligne (état activé), ce qui écarte les segments de l'opsine et permet à une molécule de protéine G (en brun) de s'intercaler. La cascade de réactions biochimiques de la phototransduction est alors engagée.

Spectre d'absorption des photorecepteurs de la rétine
Selon la nature de l'opsine on distingue quatre types de pigments rétiniens qui absorbent chacun dans une gamme de longueurs d'ondes différente. Chaque cellule photoréceptrice ne contient qu'un seul type de pigment, c'est à dire un seul type d'opsine car le rétinal est identique pour tous les pigments.
Remarque. C'est le rétinal qui absorbe les photons mais la gamme de longueurs d'ondes absorbée dépend de la nature de l'opsine.
Image modifiée : SVT 1eS, Nathan 2011 p. 352 d'après www.inrp.fr
Selon la nature de l'opsine on distingue quatre types de pigments rétiniens qui absorbent chacun dans une gamme de longueurs d'ondes différente. Chaque cellule photoréceptrice ne contient qu'un seul type de pigment, c'est à dire un seul type d'opsine car le rétinal est identique pour tous les pigments.
Remarque. C'est le rétinal qui absorbe les photons mais la gamme de longueurs d'ondes absorbée dépend de la nature de l'opsine.
Image modifiée : SVT 1eS, Nathan 2011 p. 352 d'après www.inrp.fr
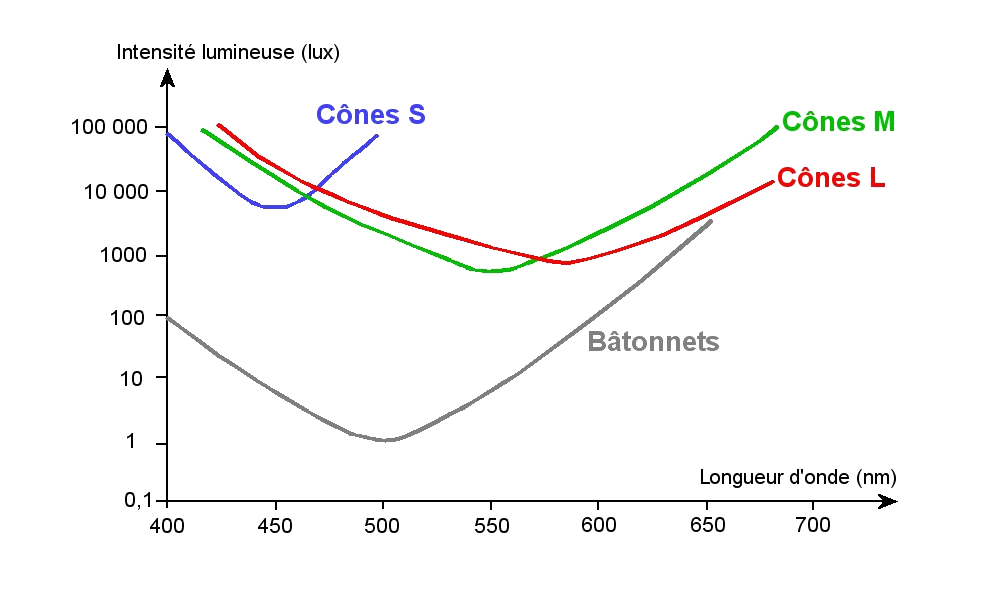
- Il existe un seul type de bâtonnet car ils contiennent tous le même pigment (la rhodopsine) qui absorbe toutes les longueurs d'onde du spectre de la lumière visible (avec cependant un maximum vers 500 nm). Les messages nerveux provenant des bâtonnets sont donc peçus en noir et blanc.
- Il existe 3 types de cônes contenant chacun un pigment différent. L'opsine S permet l'absorption dans le bleu (maximum d'absorption à 420 nm), l'opsine M permet l'absorption dans le vert (maximum d'absorption à 530 nm) et l'opsine L permet l'absorption dans le rouge (maximum d'absorption à 560 nm). On qualifie donc les cônes de S, M et L (ou bleu, vert et rouge) selon l'opsine qu'ils contiennent.
Les lettres conventionnelles S, M et L proviennent des mots anglais Short, Medium et Long wavelenght, qui correspondent respectivement aux courtes, moyennes et grandes longueurs d'ondes.
- Les spectres d'absorption des pigments se chevauchent. Pour une longueur d'onde donnée, les trois types de cônes sont stimulés, mais à différents degrés. Cela permet la perception des couleurs intermédiaires.
- Il existe 3 types de cônes contenant chacun un pigment différent. L'opsine S permet l'absorption dans le bleu (maximum d'absorption à 420 nm), l'opsine M permet l'absorption dans le vert (maximum d'absorption à 530 nm) et l'opsine L permet l'absorption dans le rouge (maximum d'absorption à 560 nm). On qualifie donc les cônes de S, M et L (ou bleu, vert et rouge) selon l'opsine qu'ils contiennent.
Les lettres conventionnelles S, M et L proviennent des mots anglais Short, Medium et Long wavelenght, qui correspondent respectivement aux courtes, moyennes et grandes longueurs d'ondes.
- Les spectres d'absorption des pigments se chevauchent. Pour une longueur d'onde donnée, les trois types de cônes sont stimulés, mais à différents degrés. Cela permet la perception des couleurs intermédiaires.
1 2
2 
1. Seuils de sensibilité des photorécepteurs - 2. Propriétés des photoréceptreurs
Les bâtonnets sont très sensibles à la lumière (1) ce qui permet la vision sous faible éclairement (seuil de sensibilité 10-14 Watts, soit la puissance d'un seul photon). La régénération de la rhodopsine (retour à l'état stable) ne s'opère qu'à l'obcurité. Ils permettent la vision crépusculaire. Si une lumière intense persiste, la rhodopsine reste activée et les bâtonnets sont saturés ne peuvent plus fournir de réponse. Les cônes prennent alors le relai (2).
Les cônes ont besoin de davantage de lumière que les bâtonnets pour être stimulés (1) et n'interviennent donc pas dans la vision nocturne (2). Par contre leurs pigments peuvent se régénérer (retour à l'état passif) à la lumière ce qui permet la vision diurne (2).
Images : 1 d'après : acces.ens-lyon.fr - 2 SVT 1S, Bordas 2011 p. 308
 2
2 
1. Seuils de sensibilité des photorécepteurs - 2. Propriétés des photoréceptreurs
Les bâtonnets sont très sensibles à la lumière (1) ce qui permet la vision sous faible éclairement (seuil de sensibilité 10-14 Watts, soit la puissance d'un seul photon). La régénération de la rhodopsine (retour à l'état stable) ne s'opère qu'à l'obcurité. Ils permettent la vision crépusculaire. Si une lumière intense persiste, la rhodopsine reste activée et les bâtonnets sont saturés ne peuvent plus fournir de réponse. Les cônes prennent alors le relai (2).
Les cônes ont besoin de davantage de lumière que les bâtonnets pour être stimulés (1) et n'interviennent donc pas dans la vision nocturne (2). Par contre leurs pigments peuvent se régénérer (retour à l'état passif) à la lumière ce qui permet la vision diurne (2).
Images : 1 d'après : acces.ens-lyon.fr - 2 SVT 1S, Bordas 2011 p. 308
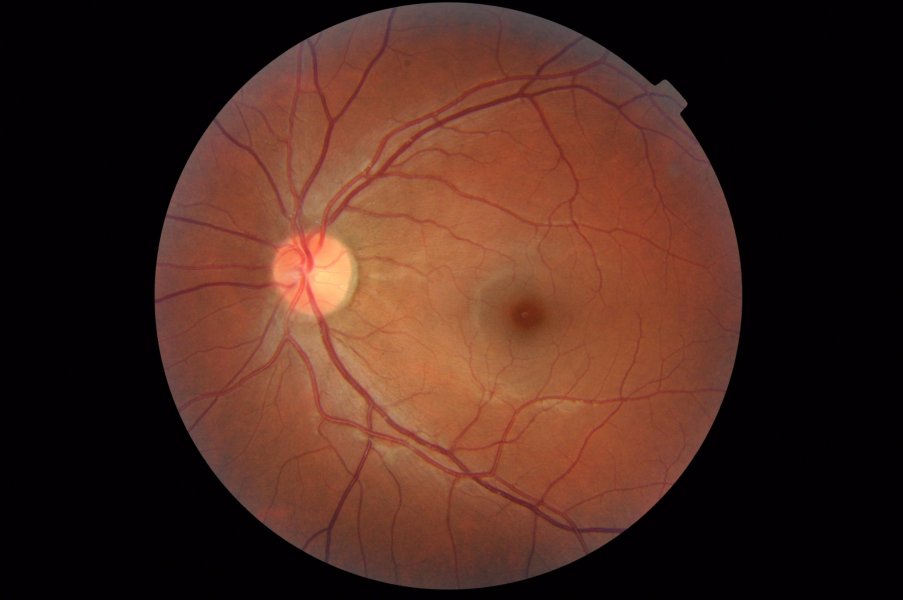
Pour chaque œil, la rétine humaine forme un disque de 42 mm de diamètre et compte environ 100 millions de bâtonnets et 3 millions de cônes. La rétine centrale ou macula (ou tache jaune) est située dans l'axe optique. Elle occupe un champ circulaire d’environ 6 mm au centre de l'image ci-contre. Les vaisseaux sanguins de la choroïde convergent en un point qui est aussi le point de départ du nerf optique.
Image : commons.wikimedia.org - OptometrusPrime
{kind=link}
Coupe de rétine au niveau de la macula (singe MP x 240)
Au centre de la macula, la fovéa forme une dépression de 200 µm de diamètre. On y observe un déplacement latéral des neurones et des fibres nerveuses. Seuls demeurent les photorécepteurs qui, du fait de la faible épaisseur de la rétine à ce niveau, reçoivent davantage de lumière. Remarquer l'abondance des cellules ganglionnaires au niveau de la macula.
S sclérotique ; C choroïde ; flèche épithélium (pigmenté chez l'Homme) ; Pn noyaux des photorécepteurs ; B noyaux des neurones bipolaires (auxquels sont associés les noyaux des cellules horizontales, côté interne, et amacrines, côté externe, qui apparaissent plus gros) ; G cellules ganglionnaires ; F fibres nerveuses.
Image (modifiée) : ecfa.unige.ch
Au centre de la macula, la fovéa forme une dépression de 200 µm de diamètre. On y observe un déplacement latéral des neurones et des fibres nerveuses. Seuls demeurent les photorécepteurs qui, du fait de la faible épaisseur de la rétine à ce niveau, reçoivent davantage de lumière. Remarquer l'abondance des cellules ganglionnaires au niveau de la macula.
S sclérotique ; C choroïde ; flèche épithélium (pigmenté chez l'Homme) ; Pn noyaux des photorécepteurs ; B noyaux des neurones bipolaires (auxquels sont associés les noyaux des cellules horizontales, côté interne, et amacrines, côté externe, qui apparaissent plus gros) ; G cellules ganglionnaires ; F fibres nerveuses.
Image (modifiée) : ecfa.unige.ch

Coupe de rétine au niveau du nerf optique (MP x 80)
Les fibres nerveuses qui tapissent la face interne de la rétine traversent les enveloppes de l'œil pour former le nerf optique. Cela entraîne l'interruption de la couche de photorécepteurs et marque le point aveugle. Remarquer l'épaisseur importante de la couche de fibres nerveuses. Sur la partie droite du cliché le décollement entre la rétine et de la choroïde est un artéfact dû à la préparation microscopique utilisée.
Image : ecfa.unige.ch
Les fibres nerveuses qui tapissent la face interne de la rétine traversent les enveloppes de l'œil pour former le nerf optique. Cela entraîne l'interruption de la couche de photorécepteurs et marque le point aveugle. Remarquer l'épaisseur importante de la couche de fibres nerveuses. Sur la partie droite du cliché le décollement entre la rétine et de la choroïde est un artéfact dû à la préparation microscopique utilisée.
Image : ecfa.unige.ch
1  2
2 
1. Surface de la rétine (MEB) à proximité de la fovéa (A) et à sa périphérie (B) - 2 Distribution spatiale des cônes et des bâtonnets sur la rétine
La fovéa contient uniquement des cônes (150 000 par mm2) dont la densité diminue très rapidement au delà. Cette zone permet la vision des couleurs le jour. Les bâtonnets sont répartis sur la rétine périphérique qui est spécialisée dans la vision en niveaux de gris sous faible éclairement. Le point aveugle est totalement dépourvu de photorécepteurs.
Images : 1 SVT 1eS, Belin 2011 p. 295 - 2 SVT 1e S Nathan 2011 p. 351
 2
2 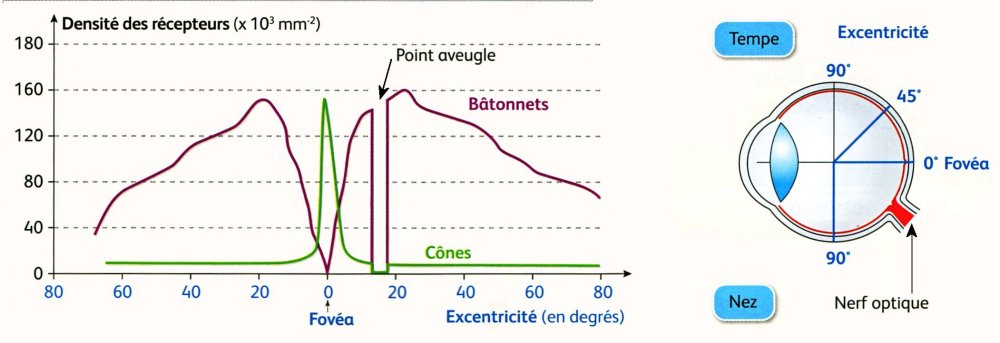
1. Surface de la rétine (MEB) à proximité de la fovéa (A) et à sa périphérie (B) - 2 Distribution spatiale des cônes et des bâtonnets sur la rétine
La fovéa contient uniquement des cônes (150 000 par mm2) dont la densité diminue très rapidement au delà. Cette zone permet la vision des couleurs le jour. Les bâtonnets sont répartis sur la rétine périphérique qui est spécialisée dans la vision en niveaux de gris sous faible éclairement. Le point aveugle est totalement dépourvu de photorécepteurs.
Images : 1 SVT 1eS, Belin 2011 p. 295 - 2 SVT 1e S Nathan 2011 p. 351
-
Pour prendre conscience du point aveugle, fermer un œil et tenir un crayon à bout de bras de telle sorte que la gomme située à l'une de ses extrtémités soit au centre du champ de vision de l'œil ouvert. Tout en continuant à regarder en face, écarter doucement le bras sur le côté extérieur. La gomme disparaît quand son image se forme sur le point aveugle (angle d'environ 10° par rapport à l'axe optique) puis réapparaît ensuite ou si on monte ou descend le crayon.
-
Pour mieux voir une étoile peu lumineuse il faut regarder à côté ce qui dirige le rayon lumineux vers la périphérie de la rétine où les bâtonnets sont abondants. Si on fixe l'étoile, son image se forme au niveau de la fovéa et son intensité lumineuse est insuffisante pour stimulet les cônes.
-
La répartition et la proportion des cônes et de bâtonnets est variable selon l'espèce. Les animaux nocturnes n'ont que des bâtonnets dans leur rétine et ne sont donc pas capables de voir les couleurs. En revanche, quelques animaux sont exclusivement diurnes (écureuils) et n'ont que des cônes.
-
Les photorécepteurs représentent 70 % des cellules réceptrices de l'organisme. Cela souligne l'importance de l'information visuelle dans la perception de l'environnement chez l'Homme.
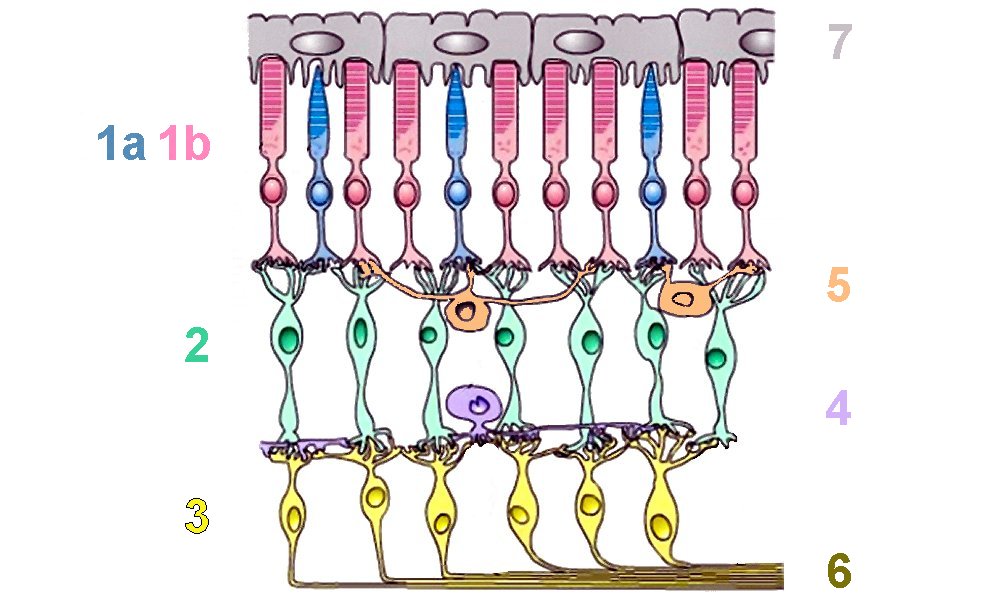
Organisation cellulaire de la rétine humaine
À côté des photorécepteurs que sont les cônes (1a) et les bâtonnets (1b) on distingue 4 autres types de neurones à fonction d'intégration : les cellules bipolaires (2), les cellules ganglionnaires (3), les cellules amacrines (4) et les cellules horizontales (5). À cela s'ajoutent les fibres du nerf optique (6), issues des cellules ganglionnaires et un épithélium (7), contenant de pigments mélaniques chez l'Homme et dans lequel sont enchâssés les photorécepteurs, qui absorbe les rayons parasites et assure le renouvellement des pigments photosensibles.
Image : acces.inrp.fr (modifiée)
À côté des photorécepteurs que sont les cônes (1a) et les bâtonnets (1b) on distingue 4 autres types de neurones à fonction d'intégration : les cellules bipolaires (2), les cellules ganglionnaires (3), les cellules amacrines (4) et les cellules horizontales (5). À cela s'ajoutent les fibres du nerf optique (6), issues des cellules ganglionnaires et un épithélium (7), contenant de pigments mélaniques chez l'Homme et dans lequel sont enchâssés les photorécepteurs, qui absorbe les rayons parasites et assure le renouvellement des pigments photosensibles.
Image : acces.inrp.fr (modifiée)
Les bâtonnets et les cônes communiquent (par contact synaptique) avec les neurones bipolaires qui, à leur tour, communiquent avec les cellules ganglionnaires. Les cellules horizontales et les cellules amacrines assurent une communication latérale (elles permettent l'intégration de l'information avant son acheminement au cerveau). Au bilan, la rétine ne rediffuse pas l'image, mais la transforme en message nerveux acheminé vers le cerveau par les fibres nerveuses issues des cellules ganglionnaires et qui forment le nerf optique.
-
Le message nerveux au sens strict ne nait véritablement qu'à partir des cellules ganglionnaires. Les mécanismes de communication entre les cellules de la rétine sont complexes et particuliers. Ils sont hors programme.
-
Le temps nécessaire pour que ces processus chimiques se réalisent est d'environ 45 ms. Cela entraîne le phénomène de persitance rétinienne qui fait que deux images se succédant rapidement apparaissent en continu (principe du cinéma et de la télévision).
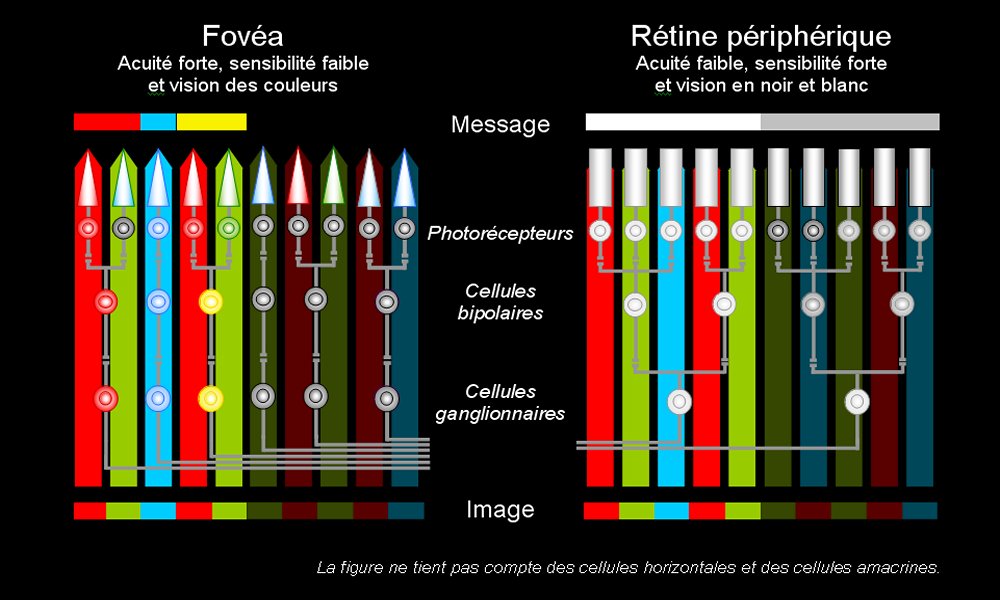
Les champs récepteurs
Chaque cellule bipolaire est reliée à plusieurs bâtonnets ou à plusieurs cônes et chaque cellule ganglionnaire reçoit des informations de plusieurs cellules bipolaires. Ainsi pour 100 millions de photorécepteurs dans chaque rétine nous possédont seulement 1 million de cellules ganglionnaires. Tous les bâtonnets et les cônes qui envoient l'information à une même cellule ganglionnaire (via les cellues bipolaires, horizontales et amacrines) forment le champ récepteur (circulaire) de cette cellule.
Chaque cellule bipolaire est reliée à plusieurs bâtonnets ou à plusieurs cônes et chaque cellule ganglionnaire reçoit des informations de plusieurs cellules bipolaires. Ainsi pour 100 millions de photorécepteurs dans chaque rétine nous possédont seulement 1 million de cellules ganglionnaires. Tous les bâtonnets et les cônes qui envoient l'information à une même cellule ganglionnaire (via les cellues bipolaires, horizontales et amacrines) forment le champ récepteur (circulaire) de cette cellule.
Les cellules ganglionnaires de la fovéa, qui contient des cônes, ont des champs récepteurs beaucoup plus petits que celles de la périphérie, contenant des bâtonnets. Cela a deux conséquences :
- l'acuité visuelle (capacité à séparer deux points) est améliorée dans la fovéa car chaque fibre nerveuse du nerf optique concerne une petite surface de réception photosensible ;
- la sensibilité de la rétine périphérique est améliorée de nuit car les stimulations de chacun des bâtonnets reliés à une même fibre nerveuse s'additionnent. On parle de sommation (spatiale).
- l'acuité visuelle (capacité à séparer deux points) est améliorée dans la fovéa car chaque fibre nerveuse du nerf optique concerne une petite surface de réception photosensible ;
- la sensibilité de la rétine périphérique est améliorée de nuit car les stimulations de chacun des bâtonnets reliés à une même fibre nerveuse s'additionnent. On parle de sommation (spatiale).
-
Il y a davantage de cellules ganglionnaires au niveau de la fovéa (voir II.C ).
-
Le pouvoir séparateur est la valeur limite de l'angle permettant de séparer deux points. Chez l'Homme il est de 1 minute d'angle. Pour des raisons de commodité le pouvoir séparateur est quantifié par son inverse : l'acuité visuelle. Chez l'Homme l'acuité visuelle moyenne est de 10/10e ce qui correspond à la séparation de deux points sur la rétine d'environ 5 µm (il est donc possible d'avoir une acuité visuelle supérieure à 10/10e ).
-
Un faucon possède 1 000 000 de cellules par mm2 de fovéa, son acuité visuelle est 8 fois supérieure à la nôtre. Il possède de plus deux fovéas dans chaque œil. L'une reçoit la lumière de front, l'autre la reçoit de dessous. En vol, le faucon voit en même temps ce qui se passe devant lui et ce qui se passe au dessous. Il peut ainsi diriger sont vol et repérer en même temps une proie au sol.
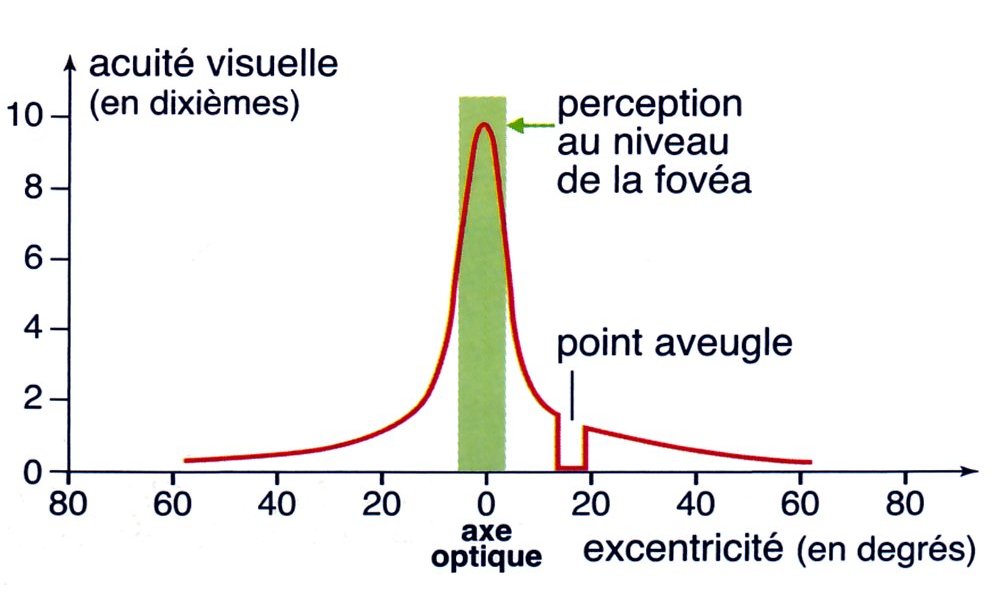
Acuité visuelle de la rétine
L'acuité visuelle est maximale au niveau de la fovéa (à condition que l'éclairement soit suffisant). C'est la raison pour laquelle, lors d'une activité de précision comme la lecture, le sujet recherche le bon éclairement et bouge sans arrêt les yeux de manière à diriger les fovéas des deux yeux vers les caractères regardés.
Image : SVT 1eS, Bordas 2011, p. 308
L'acuité visuelle est maximale au niveau de la fovéa (à condition que l'éclairement soit suffisant). C'est la raison pour laquelle, lors d'une activité de précision comme la lecture, le sujet recherche le bon éclairement et bouge sans arrêt les yeux de manière à diriger les fovéas des deux yeux vers les caractères regardés.
Image : SVT 1eS, Bordas 2011, p. 308

Le daltonisme
Le daltonisme (= dyschromatopsie découverte par John Dalton (lien externe) ) se manifeste par des perturbations de la vision des couleurs. Il est lié à une ou plusieurs mutations sur les gènes des opsines ce qui entraîne la défaillance dans la sensibilité à des longueurs d'ondes particulières. La forme la plus fréquente résulte de la déficience plus ou moins complète en cônes verts (deutéranopie). Plus rarement la mutation affecte les cônes rouges (protanopie) ou bleus (tritanopie) voire les trois (achromatopsie). Il s'agit d'un trouble héréditaire qui affecte beaucoup plus souvent les hommes (8%) que les femmes (0,5%) car c'est un caractère lié au sexe (gène porté par le chromosome X).
Image : SVT 1eS, Nathan 2011 p. 353
Le daltonisme (= dyschromatopsie découverte par John Dalton (lien externe) ) se manifeste par des perturbations de la vision des couleurs. Il est lié à une ou plusieurs mutations sur les gènes des opsines ce qui entraîne la défaillance dans la sensibilité à des longueurs d'ondes particulières. La forme la plus fréquente résulte de la déficience plus ou moins complète en cônes verts (deutéranopie). Plus rarement la mutation affecte les cônes rouges (protanopie) ou bleus (tritanopie) voire les trois (achromatopsie). Il s'agit d'un trouble héréditaire qui affecte beaucoup plus souvent les hommes (8%) que les femmes (0,5%) car c'est un caractère lié au sexe (gène porté par le chromosome X).
Image : SVT 1eS, Nathan 2011 p. 353

Le test d'Ishihara
C'est le plus connu des tests de vision des couleurs permettant de repérer le daltonisme. Chaque planche est constituée d'une mosaïque de points de couleurs différentes à plusieurs degrés de taille, de saturation et de luminosité, au sein de laquelle apparaît une forme reconnaisable.
Image : www.daltoniens.fr
Que représente chacune des 6 images ? Noter les résultats puis consulter l'interprétation du test d'Ishiara.
C'est le plus connu des tests de vision des couleurs permettant de repérer le daltonisme. Chaque planche est constituée d'une mosaïque de points de couleurs différentes à plusieurs degrés de taille, de saturation et de luminosité, au sein de laquelle apparaît une forme reconnaisable.
Image : www.daltoniens.fr
Que représente chacune des 6 images ? Noter les résultats puis consulter l'interprétation du test d'Ishiara.

L'héméralopie est une cécité nocturne. Elle est génértalement due à une carence en vitamine A qui entaîne un déficit en rétinal et donc en pigments visuels. Les cônes peuvent continuer à fonctionner mais les bâtonnets cessent leur activité, ce qui entraîne une cécité nocturne (ou héméralopie).
La vitamine A est une vitamine liposoluble notamment présente dans les lipides d'origine animale et le foie. Elle est également présente certains végétaux comme carotte (carotène), l'oseille, les épinards, le navet ou la mangue...
Image : cashenka.skyrock.com
La vitamine A est une vitamine liposoluble notamment présente dans les lipides d'origine animale et le foie. Elle est également présente certains végétaux comme carotte (carotène), l'oseille, les épinards, le navet ou la mangue...
Image : cashenka.skyrock.com
1  2
2 
1. Rétinite pigmentaire : vision en tunnel (forme avancé de la maladie)
2. Rétine d'une personne atteinte de rétinite pigmentaire
Images : 1 www.ophtalmo.net - 2 www.larousse.fr
La rétinite pigmentaire est une maladie génétique impliquant une quarantaine de gènes portés par les chromosomes 8, 13 et X. Elle débute par une héméralopie puis, progressivement, le champ visuel se rétrécit donnant une impression de « vision en tunnel » (1). Cette altération est due à la destruction progressive des bâtonnets puis des cônes, ce qui a pour conséquence l’apparition progressive de petits dépôts pigmentés (2) dans la rétine donnant ainsi le nom à la maladie.
On observe souvent une modification de la rhodopsine dans les bâtonnets de l’œil. Contrairement à ce qui se passe dans un œil sain dans lequel elle est régulièrement renouvelée dans sa totalité, lors d’une rétinite pigmentaire des fragments de « l’ancienne » molécule subsistent et ils détériorent progressivement les cellules visuelles.
Source : www.psi.ch
 2
2 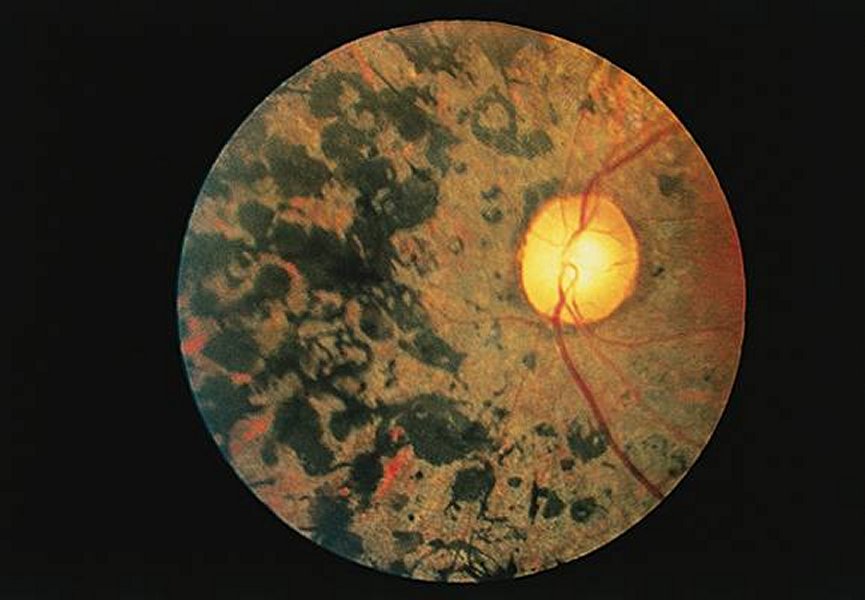
1. Rétinite pigmentaire : vision en tunnel (forme avancé de la maladie)
2. Rétine d'une personne atteinte de rétinite pigmentaire
Images : 1 www.ophtalmo.net - 2 www.larousse.fr
La rétinite pigmentaire est une maladie génétique impliquant une quarantaine de gènes portés par les chromosomes 8, 13 et X. Elle débute par une héméralopie puis, progressivement, le champ visuel se rétrécit donnant une impression de « vision en tunnel » (1). Cette altération est due à la destruction progressive des bâtonnets puis des cônes, ce qui a pour conséquence l’apparition progressive de petits dépôts pigmentés (2) dans la rétine donnant ainsi le nom à la maladie.
On observe souvent une modification de la rhodopsine dans les bâtonnets de l’œil. Contrairement à ce qui se passe dans un œil sain dans lequel elle est régulièrement renouvelée dans sa totalité, lors d’une rétinite pigmentaire des fragments de « l’ancienne » molécule subsistent et ils détériorent progressivement les cellules visuelles.
Source : www.psi.ch
1  2
2 
1. Dégénérescence maculaire liée à l'âge (DMLA) : effets de la maladie
2. Rétine d'une personne atteinte de DMLA
La DMLA entraîne une perte progressive de la vision centrale (1), qui devient de plus en plus floue. C'est une dégénérescence progressive des cellules de la macula qui ne provoque jamais de cécité totale. Elle touche 12 % de la population entre 65 et 75 ans.
Images 1 www.ophtalmologie-jmschepens.com - 2 www.usherbrooke.ca
 2
2 
1. Dégénérescence maculaire liée à l'âge (DMLA) : effets de la maladie
2. Rétine d'une personne atteinte de DMLA
La DMLA entraîne une perte progressive de la vision centrale (1), qui devient de plus en plus floue. C'est une dégénérescence progressive des cellules de la macula qui ne provoque jamais de cécité totale. Elle touche 12 % de la population entre 65 et 75 ans.
Images 1 www.ophtalmologie-jmschepens.com - 2 www.usherbrooke.ca
Organisation cellulaire de la rétine humaine● La rétine est une membrane qui tapisse le fond de l'œil et sur laquelle se forme l'image d'un objet. De la face externe à la face interne elle est formée de trois couches principales :
- une couche de récepteurs sensoriels de la vision appelés photorécepteurs (cônes et bâtonnets) qui contiennent un pigment rétinien protéique, l'opsine (associé au rétinal) ;
- une couche de cellules bipolaires (auxquelles s'ajoutent le des cellules horizontales et des cellules amacrines) ;
- une couche de cellules ganglionnaires à l'origine des fibres du nerf optique qui achemine au cerveau le message nerveux généré par le stimulus visuel.
● Les bâtonnets sont situés dans la rétine périphérique et sont sensibles aux faibles intensités lumineuses (vision crépusculaire uniquement). Ils contiennent tous la même opsine (rhodopsine) qui absorbe toutes les longueurs d'onde du spectre de la lumière visible et n'interviennent pas dans la vision des couleurs. Les cônes sont concentrés dans l'axe optique de l'œil (fovéa) et ont un seuil de stimulation élevé (vision diurne uniquement). La rétine humaine possède trois types de cônes respectivement sensibles au bleu (opsine S), au vert (opsine M) et au rouge (opsine L). Cela permet la vision des couleurs (trichromate).
● Le daltonisme se manifeste par des perturbations de la vision des couleurs. Il est lié à une ou plusieurs mutations sur les gènes des pigments rétiniens des cônes ce qui entraîne la défaillance dans la sensibilité à des longueurs d'ondes particulières. Cela se traduit par des perturbations de la vision des couleurs.

Localisation chromosomique des gènes des pigments rétiniens
Chacune des 4 opsines est codée par un gène différent. Le gène codant l'opsine de la rhodopsine des bâtonnets (Rho) est situé sur le chromosme 3, celui de l'opsine du pigment absorbant le bleu (cônes S) est situé sur le chromosome 7 tandis que les gènes codant les opsines absorbant le rouge (cônes L) et le vert (cônes M) sont étroitement liés sur le chromosome X.
Chacune des 4 opsines est codée par un gène différent. Le gène codant l'opsine de la rhodopsine des bâtonnets (Rho) est situé sur le chromosme 3, celui de l'opsine du pigment absorbant le bleu (cônes S) est situé sur le chromosome 7 tandis que les gènes codant les opsines absorbant le rouge (cônes L) et le vert (cônes M) sont étroitement liés sur le chromosome X.
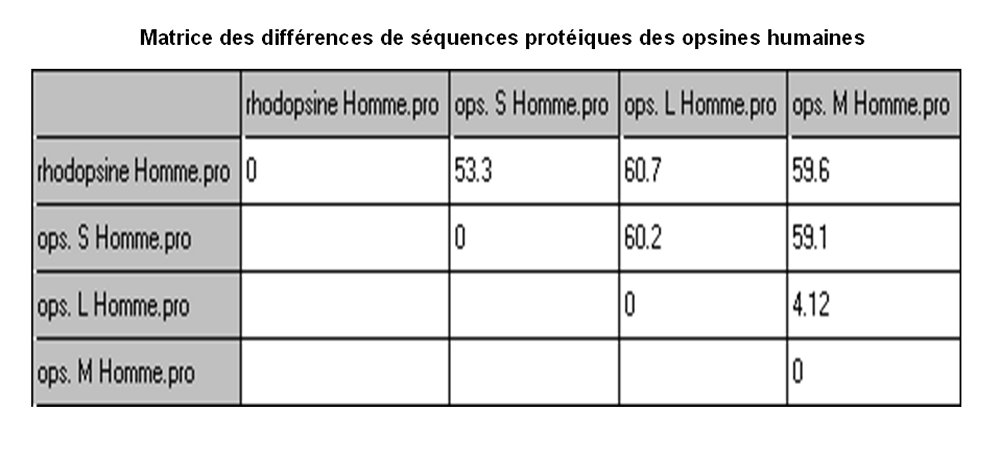
Comparaison des séquences d'acides aminés des pigments rétiniens
Matrice des différences (en pourcentages). C'est un tableau à double entrée indiquant ici le pourcentage de différences entre les opsines prises deux à deux.
Les opsines absorbant le rouge ou le vert, formées de 364 acides aminés, ont une séquence identique à plus de 95%. Chacune partage plus de 40% de ses acides aminés avec l'opsine absorbant le bleu (formée de 348 acides aminés) ou avec la rhodopsine des bâtonnets.
Or, on considère qu'une similitude entre deux protéines, supérieure à 20% (ou supérieure à 40% entre deux gènes), ce qui est le cas des opsines, indique une origine commune. On parle alors de famille multigénique.
Figure construite à l'aide du logiciel Phylogène.
Matrice des différences (en pourcentages). C'est un tableau à double entrée indiquant ici le pourcentage de différences entre les opsines prises deux à deux.
Les opsines absorbant le rouge ou le vert, formées de 364 acides aminés, ont une séquence identique à plus de 95%. Chacune partage plus de 40% de ses acides aminés avec l'opsine absorbant le bleu (formée de 348 acides aminés) ou avec la rhodopsine des bâtonnets.
Or, on considère qu'une similitude entre deux protéines, supérieure à 20% (ou supérieure à 40% entre deux gènes), ce qui est le cas des opsines, indique une origine commune. On parle alors de famille multigénique.
Figure construite à l'aide du logiciel Phylogène.

Mécanismes génétique à l'origine d'une famille multigénique
Un gène ancestral (A) subit des duplications (A'), des transpositions (déplacement sur un autre chromosome) et des mutations géniques (modifications de séquence (B)) qui peuvent intervenir avant ou après transposition).
Plus une duplication est ancienne et plus les mutations géniques ont pu s'accumuler indépendamment sur les copies. Il en résulte que les protéines codées restent apparentées (au moins 20% d'acides aminés en commun) mais peuvent remplir des fonctions différentes. On ne parle de famille multigénique.
Un gène ancestral (A) subit des duplications (A'), des transpositions (déplacement sur un autre chromosome) et des mutations géniques (modifications de séquence (B)) qui peuvent intervenir avant ou après transposition).
Plus une duplication est ancienne et plus les mutations géniques ont pu s'accumuler indépendamment sur les copies. Il en résulte que les protéines codées restent apparentées (au moins 20% d'acides aminés en commun) mais peuvent remplir des fonctions différentes. On ne parle de famille multigénique.
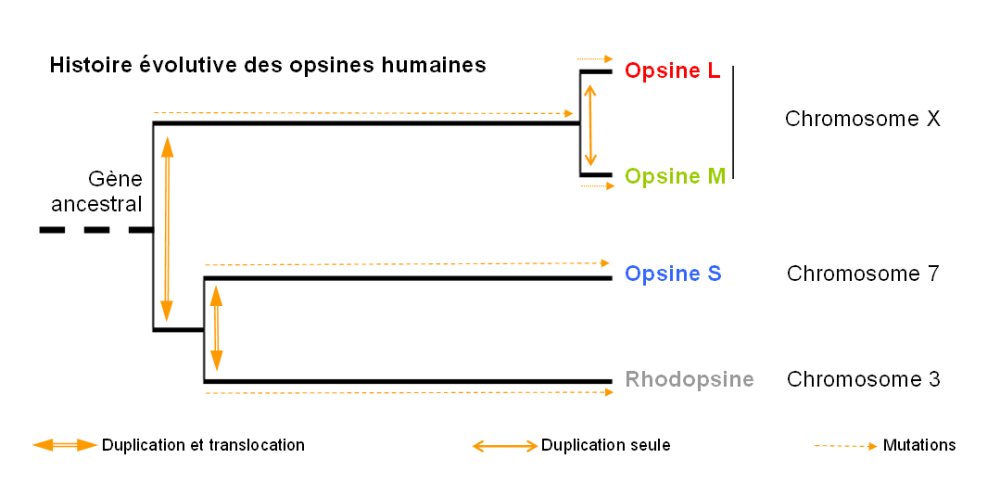
La famille multigénique des opsines
- Une première duplication avec translocation et mutations géniques affecte le gène ancestral et permet d'obtenir le gène à l'origine de la rhodopsine et de l'opsine S d'une part et le gène à l'origine des opsines rouge et verte (situé sur le chromosome X).
- Une deuxième duplication avec translocation et mutations affecte le gène de la rhodopsine et permet d'obtenir le gène de la rhodopsine (chromosome 3) et le gène de l'opsine S (chromosome 7).
- Une troisième duplication suivie de mutations ponctuelles (mais sans translocation) affecte le le gène situé sur le chromosome X et permet d'obtenir le gène de l'opsine M et de l'opsine L.
Figure réalisée à partir de l'exploitation des données fournies par le logiciel Phylogène.
- Une première duplication avec translocation et mutations géniques affecte le gène ancestral et permet d'obtenir le gène à l'origine de la rhodopsine et de l'opsine S d'une part et le gène à l'origine des opsines rouge et verte (situé sur le chromosome X).
- Une deuxième duplication avec translocation et mutations affecte le gène de la rhodopsine et permet d'obtenir le gène de la rhodopsine (chromosome 3) et le gène de l'opsine S (chromosome 7).
- Une troisième duplication suivie de mutations ponctuelles (mais sans translocation) affecte le le gène situé sur le chromosome X et permet d'obtenir le gène de l'opsine M et de l'opsine L.
Figure réalisée à partir de l'exploitation des données fournies par le logiciel Phylogène.
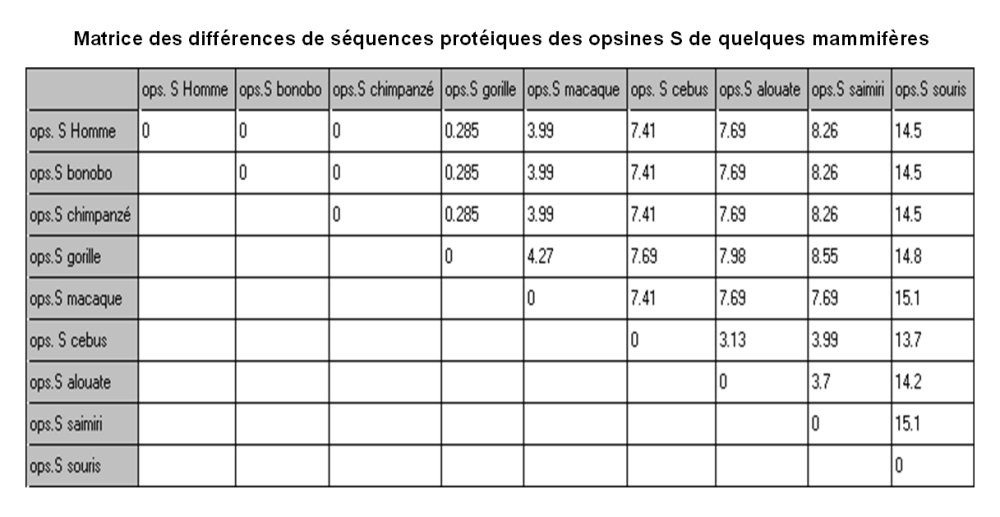
Comparaison de la séquence de l'opsine S chez quelques primates
Matrice des différences (en pourcentages). C'est un tableau à double entrée indiquant ici le nombre de différences en pourcentage dans la séquence des acides aminés de l'opsine S pour les espèces prises deux à deux.
Figure construite à l'aide du logiciel Phylogène.
Matrice des différences (en pourcentages). C'est un tableau à double entrée indiquant ici le nombre de différences en pourcentage dans la séquence des acides aminés de l'opsine S pour les espèces prises deux à deux.
Figure construite à l'aide du logiciel Phylogène.
Principe de construction d'une arbre de parenté à partir de données moléculaires
- Deux espèces qui présentent une même nouveauté évolutive l'ont hérité d'un ancêtre commun chez qui l'innovation est apparue.
- Pour deux espèces données, possédant une même protéine (protéines homologues), plus le nombre de différences dans la séquence des acides aminés est important, plus le nombre de mutations du gène codant cette protéine est important, plus l'ancêtre commun aux deux espèces est éloigné dans le temps.
Dans un arbre de parenté (= arbre phylogénétique) chaque nœud correspond à un ancêtre commun et chaque extrémité de branche à un organisme (actuel ou fossile).
- Deux espèces qui présentent une même nouveauté évolutive l'ont hérité d'un ancêtre commun chez qui l'innovation est apparue.
- Pour deux espèces données, possédant une même protéine (protéines homologues), plus le nombre de différences dans la séquence des acides aminés est important, plus le nombre de mutations du gène codant cette protéine est important, plus l'ancêtre commun aux deux espèces est éloigné dans le temps.
Dans un arbre de parenté (= arbre phylogénétique) chaque nœud correspond à un ancêtre commun et chaque extrémité de branche à un organisme (actuel ou fossile).
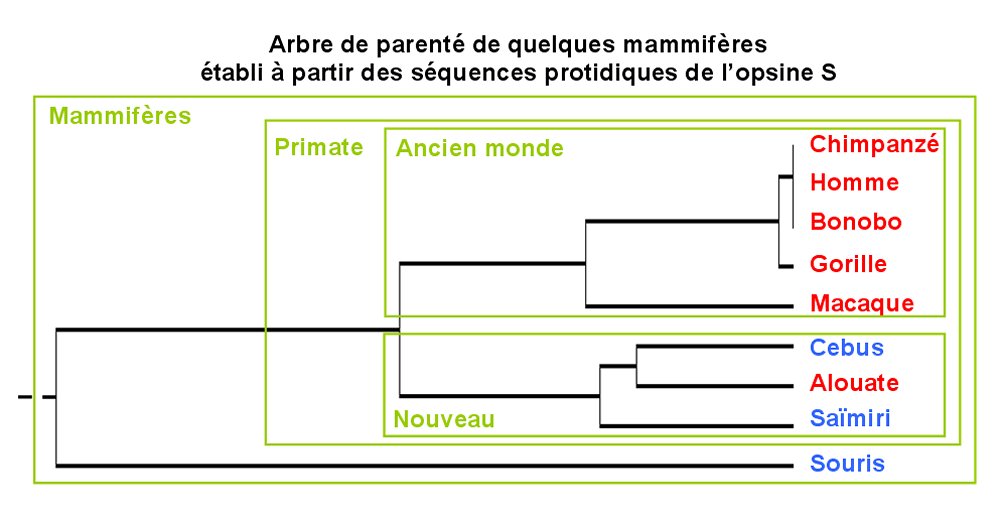
L'Homme, le Chimpanzé et le Bonobo sont le plus étroitement apparentés car ils ont l'ancêtre commun le plus récent. L'Homme est plus proche des singes de l'ancien monde (Afrique, Asie et Europe) que de ceux du nouveau monde (Amérique).
Figure construite à l'aide du logiciel Phylogène.
1.  2
2 
1. L ocalisation chromosomique des gènes des pigments rétiniens
2. Les opsines S,M et L et vision des couleurs chez les primates
L'Homme et les singes de l'ancien monde (Afrique, Asie et Europe) possèdent trois gènes différents (1) codant trois opsines différentes responsables de la vision des couleurs : S (sur le chromosome 7) ; M et L (sur le chromosome X). Ils sont trichromates. Les singes du nouveau monde (Amérique) possèdent le gène S (sur le chromosome 7) et un seul gène (sur le chromosome X) dont il existe plusieurs allèles codant pour une opsine qui absorbe soit dans le rouge soit dans le vert. Ils sont dichromates comme la plupart des mammifères.
Remarque. Les mâles (XY) des singes du nouveau monde sont forcément dichromates, mais les femelles (XX) peuvent être trichromates si elles possèdent deux allèles différents sur chacun de leurs deux chromosomes X, l'un codant une opsine absorbant dans le rouge et l'autre codant une opsine absobant dans le vert.
Image : SVT 1S, Belin 2011 p. 298 .
2 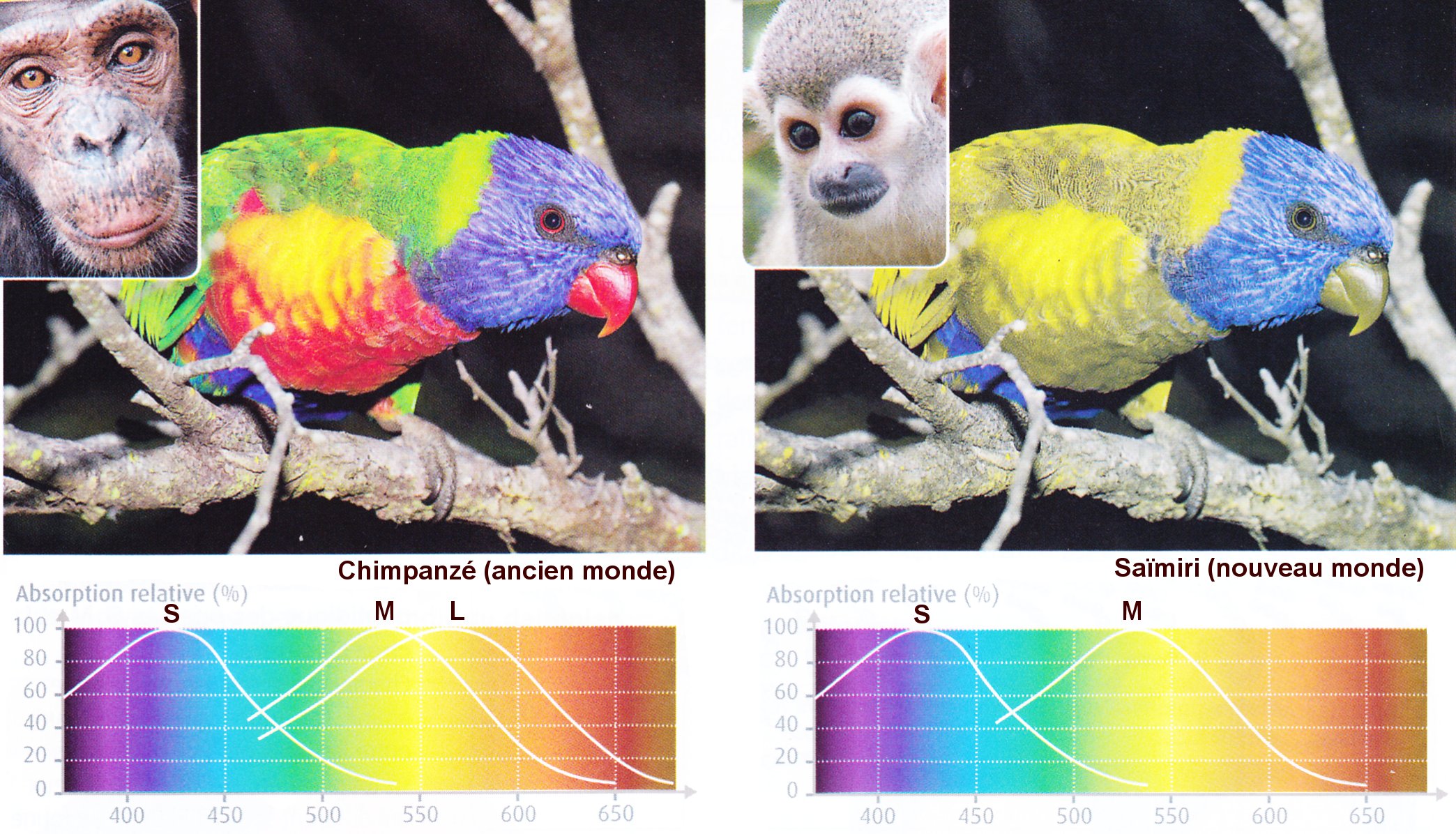
1. L ocalisation chromosomique des gènes des pigments rétiniens
2. Les opsines S,M et L et vision des couleurs chez les primates
L'Homme et les singes de l'ancien monde (Afrique, Asie et Europe) possèdent trois gènes différents (1) codant trois opsines différentes responsables de la vision des couleurs : S (sur le chromosome 7) ; M et L (sur le chromosome X). Ils sont trichromates. Les singes du nouveau monde (Amérique) possèdent le gène S (sur le chromosome 7) et un seul gène (sur le chromosome X) dont il existe plusieurs allèles codant pour une opsine qui absorbe soit dans le rouge soit dans le vert. Ils sont dichromates comme la plupart des mammifères.
Remarque. Les mâles (XY) des singes du nouveau monde sont forcément dichromates, mais les femelles (XX) peuvent être trichromates si elles possèdent deux allèles différents sur chacun de leurs deux chromosomes X, l'un codant une opsine absorbant dans le rouge et l'autre codant une opsine absobant dans le vert.
Image : SVT 1S, Belin 2011 p. 298 .
● Les gènes des opsines sont portés par les chromosomes 3 (opsine des bâtonnets), 7 (opsine S) et X (opsines M et L). Ils constituent une famille multigénique car ils dérivent tous d'un même gène ancestral par duplications géniques, transpositions et mutations indépendantes successives. Cela explique qu'ils aient conservé une séquence de nucléotides voisine et donc que la séquence des acides aminés des différentes opsines le soit également (plus de 20% de similitudes).
 Arbre de parenté des Primates
Arbre de parenté des Primates● Comme l'Homme, les singes de l'ancien monde (d'Afrique et d'Asie) sont trichromates car ils possèdent les opsines S, M et L. Ceux du nouveau monde (Amérique) sont dichromates car ils ne possèdent que deux opsines permettant la vision des couleurs (l'une sensible au bleu et l'autre sensible soit au rouge, soit au vert). Quand des espèces partagent une même nouveauté évolutive elles l'ont hérité d'un ancêtre commun qui leur est propre. L'ancêtre commun à l'Homme et aux singes de l'ancien monde est donc plus récent que leur ancêtre commun avec les singes du nouveau monde avec qui nous partageons d'autres nouveautés évolutives des primates.
-
De la même manière, plus le nombre de différences dans la séquence des gènes des opsines (ou dans celle de ses acides aminés) est faible pour deux espèces plus elles sont apparentées. Cela permet de préciser la place de l'Homme parmi les primates trichromates. L'Homme est ainsi plus proche du Chimpanzé que du Gorille, lui même plus proche de l'Homme que le Macaque.
BILAN
● Grâce au cristallin, l'œil focalise l'image sur la rétine. Des anomalies du cristallin expliquent certains défauts de la vision.
● La rétine possède des photorécepteurs, sensibles à l'intensité lumineuse et aux trois couleurs primaires, ce qui permet la transduction du stimulus lumineux en message nerveux conduit au cerveau par le nerf optique. Des anomalies des pigments rétiniens perturbent la vision des couleurs.
● Les gènes de pigments rétiniens constituent une famille multigénique dont l'étude permet de situer l'Homme parmi les primates.
● Grâce au cristallin, l'œil focalise l'image sur la rétine. Des anomalies du cristallin expliquent certains défauts de la vision.
● La rétine possède des photorécepteurs, sensibles à l'intensité lumineuse et aux trois couleurs primaires, ce qui permet la transduction du stimulus lumineux en message nerveux conduit au cerveau par le nerf optique. Des anomalies des pigments rétiniens perturbent la vision des couleurs.
● Les gènes de pigments rétiniens constituent une famille multigénique dont l'étude permet de situer l'Homme parmi les primates.
a. Tout le monde voit 12.
b. Vision normale : 8 - Déficience rouge-vert : 3.
c. Vision normale : 5 - Déficience rouge-vert : 2.
d. Vision normale : 2 - La plupart des dischromates ne voient rien, ou de façon erronée.
e. Les sujets normaux et les dischromates très faiblement atteints ne perçoivent rien - Déficience rouge-vert : 5.
f. Vision normale : 26 - Protanopie : 6. Deutéranopie : 2.
Dichromates : absence de récepteurs au vert (vision dichromatique et non trichromatique)
Protanotpes : sensibilité au rouge diminuée.
Deutéranopes : sensibilité au vert diminuée.
Test d'Ishihara complet (lien externe) et autres tests.
Retour

L'œil et la vision (Perez) (gratuit)
De Visu (INRP) (gratuit)
Sitographie
La vision (Acces INRP)
L'oeil (SNV Jussieu)
Bibliographie (nivau universitaire)
Manuel physio humaine De Boeck
Physiologie des animaux, tome 2, Richard, Nathan U 1998
Biologie, Campbell, De Boeck 1995
Le monde du vivant, Purves, Flammarion Sc 1994
Physiologie humaine, Vander 1995
Biologie et physiologie animales, Wehner-Gehring 1999
Pistes du programme
Les traitements des déficiences du cristallin.
Le daltonisme ; la vision des couleurs chez les vertébrés.